




서 론
효모(Saccharomyces cerevisiae)는 GRAS(generally recognized as safe) 인증을 받은 산업용 미생물로 오랜 기간 빵, 맥주, 와인 등 다양한 발효식품 제조에 활용되어 왔다. 효모는 배양이 용이하고 생산 비용이 낮으며, 유전자 조작이 용이하다는 장점으로 다양한 생리활성 물질 생산에 폭넓게 이용된다[1,2].
효모 세포벽은 세포 건조중량의 약 15%–30%를 차지하며, 베타글루칸, 키틴, 만난, 단백질 등으로 구성된다[3,4]. 구성 비율은 균주, 배양 단계, 생리적 상태에 따라 달라지며, 특히 글루칸과 만난은 세포벽의 구조적 안정성과 기능적 특성을 결정하는 주요 성분으로, 식품·사료·제약 산업 등에서 고부가가치 소재로 주목받고 있다[5].
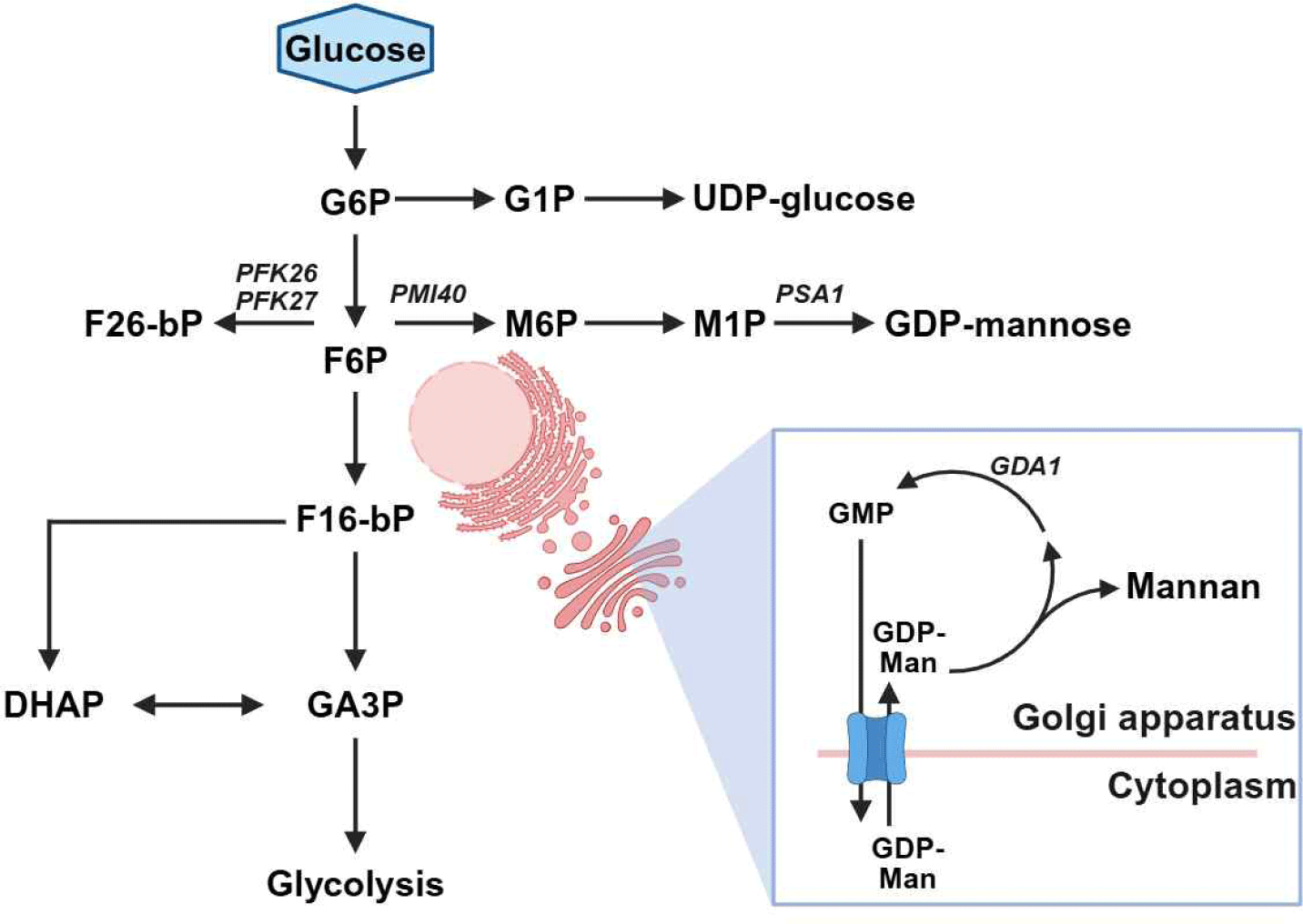
효모 유래 만난은 세포벽 질량의 약 30%–50%를 차지하는 α-결합 다당류로, α-1,6 결합을 주쇄로 하고 α-1,2 및 α-1,3 결합을 가지사슬로 갖는 구조를 지닌다[6,7]. 이러한 구조적 특성은 용해도와 생리활성에 영향을 미치며, 항산화 및 면역조절 기능을 통해 항생제 대체제 등으로 활용되고 있다[8–11]. 효모 유래 만난 생합성은 약 1,200개의 유전자에 의해 조절되며[6], 특히 fructose-6- F6P(phosphate)에서 M6P(mannose-6-phosphate), M1P(mannose-1-phosphate), GDP-mannose로 이어지는 대사 흐름이 핵심 역할을 한다(Fig. 1). PMI40 유전자는 F6P를 M6P로 전환하는 phosphomannose isomerase를, PSA1 유전자는 GDP-mannose pyrophosphorylase를 암호화하여 만난 생합성 전구체 공급을 조절한다. 생성된 GDP-mannose는 소포체를 거쳐 dolichol phosphate-mannose로 전환되어 세포벽 만난 및 단백질 글리코실화에 사용된다[5,12]. 또한, 탄소 대사 조절 유전자인 PFK26과 PFK27은 6-phosphofructo-2-kinase를 암호화하여 F2,6BP(fructose-2,6-bisphosphate) 합성을 조절하고, F2,6BP는 PFK1 활성 촉진 및 FBPase 억제를 통해 해당·당신생 합성을 전역적으로 조절하는 핵심 대사 조절 인자로 알려져 있다[13,14]. 따라서 PFK26/PFK27 결손은 해당과정 흐름을 감소시키고, 상대적으로 다른 탄소 대사 경로로의 유입을 증가시킬 수 있다. GDA1은 골지체 내 Gda1p(lumenal GDPase)를 암호화하며, GDP를 GMP(guanosine monophosphate)로 가수분해해 GMP/GDP-mannose antiporter를 구동함으로써 GDP-mannose 수송을 유지하고 단백질·지질의 만노실화 및 세포벽 구성 다당류 생합성에 필수적인 역할을 하는 것으로 알려져 있다[15,16].
효모 만난 함량은 균주에 따라 큰 차이를 보이며, 산업·의약 분야에서는 이러한 균주 간 차이를 활용한 기능성 소재 개발이 활발히 이루어지고 있다[17]. 특히 S. boulardii는 S. cerevisiae와 비교했을 때 세포벽 다당류 중 만난 비율이 더 높고, 더 두꺼운 세포벽 구조를 가지고 있다[18]. 이러한 고만난 세포벽 구조는 소화관 환경에서의 내산성, 장내 부착 억제, 면역조절과 같은 특유의 프로바이오틱 활성을 뒷받침하는 중요한 구조적 특징으로 여겨진다[19,20].
그러나 S. cerevisiae 균주 간에도 만난 축적능이 균일하지 않고, 배양 조건 변화에 따라 세포벽 조성이 크게 달라지는 것으로 알려져 있어, 균주 특이적 만난 함량을 신뢰성 있게 비교·평가할 수 있는 정량 플랫폼이 요구된다[18]. 이러한 다양한 균주에 적용 가능한 재현성 높은 만난 정량 플랫폼 구축은 기능성 효모 선발, 대사공학 균주 개발, 프로바이오틱 바이오소재 평가에 필수적이다[19].
효모 세포벽 만난의 정량 방법은 그동안 여러 방법으로 시도되어 왔다. 페놀–황산법이나 안트론법과 같은 총당 정량법은 절차가 간단하다는 장점이 있으나, 세포벽 내 다른 다당(β-glucan, 키틴 등)과 단백질의 간섭으로 인해 만난에 대한 정량 정확도가 떨어진다[5,6,11]. 또한 HPAEC–PAD(high- performance anion exchange chromatography with pulsed amperometric detection)나 GC–MS(gas chromatography-mass spectrometry) 기반 분석법은 높은 분리능을 제공하지만, 탈염 또는 유도체화 과정이 요구되어 전처리에만 수 시간에서 하루 이상이 소요되며, 시료 한 건당 분석 시간 또한 길다. 이로 인해 시료 수가 증가할수록 분석 소요 시간이 누적되어, 다수의 균주를 동일 조건에서 반복적으로 비교·분석해야 하는 고처리량 스크리닝 적용에는 한계가 있다[21–23](Table 1). 이러한 기존의 HPAEC–PAD 및 GC–MS 기반 분석법이 세포벽 다당류의 단당 조성이나 구조적 특성 분석에 중점을 둔다면, 본 연구에서 제시하는 분석법은 효모 세포벽 내 총 만난 함량을 신속하고 반복적으로 비교·평가하는 데 목적이 있다. 이에 본 연구에서는 방법 간의 직접적인 비교보다는, 표준물질 및 생물학적 시료를 이용한 내부 검증을 통해 분석법의 정확도와 재현성을 평가하였다.
이러한 한계점을 극복하기 위해 본 연구에서는 효모 세포 파쇄를 위한 균질기(homogenizer)를 적용하고, 파쇄된 효모 세포벽 만난을 산 가수분해를 통해 만노스로 전환한 후 HPLC–RID(high- performance liquid chromatography with refractive index detection) 시스템을 이용하여 정량하는 신속하고 재현성 높은 분석법을 확립하였다. 또한 대사공학적으로 만난 함량이 상이한 효모 균주를 구축하여 본 분석법의 적용성을 검증하였으며, 만난 함량이 높은 것으로 알려진 S. boulardii에서도 동일한 방법의 유효성을 확인하였다.
재료 및 방법
Escherichia coli DH5α는 S. cerevisiae 유전자 조작을 위해 필요한 플라스미드 구축을 위한 균주로 사용하였다. S. cerevisiae D452-2를 모균주로 PSA1과 PMI40 유전자를 과발현하고, PFK26과 PFK27 유전자를 결손시켜 만난 고생산 균주를 구축하였다. 또한 GDA1 유전자 결손을 통해 만난 저생산 균주를 구축하였다. S. boulardii 야생형 균주는 ATCC MYA-796에서 분양받아 사용하였다. S. boulardii 야생형 균주를 모균주로 하여 GDA1 유전자를 결손시켜 만난 저생산 균주를 구축하였다. 본 연구에서 사용한 효모 균주 목록은 Table 2에 정리하였다.
| Strains | Description | References |
|---|---|---|
| S.C | Saccharomyces cerevisiae D452-2 (MATα, leu2, his3, ura3, and can1) | [38] |
| S.C-HM | S. cerevisiae D452-2 pfk26Δ, pfk27Δ overexpressing PMI40 and PSA1 gene | In this study |
| S.C-LM | S. cerevisiae D452-2, gda1Δ | In this study |
| S.B | S. boulardii, ATCC® MYA-796TM | ATCC |
| S.B-LM | S. boulardii, gda1Δ | In this study |
E. coli는 50 μg/mL ampicillin을 첨가한 Luria-Bertani(LB) 배지(10 g/L tryptone, 5 g/L yeast extract, 10 g/L NaCl)에서 배양하였다. S. cerevisiae와 S. boulardii 균주는 20 g/L 포도당을 첨가한 YP 배지(10 g/L yeast extract, 20 g/L Bacto peptone)에 접종하여, 30℃, 1.75×g 조건에서 배양하였다. CRISPR-Cas9을 이용한 유전자 재조합 균주 선별 시에는 20 g/L 포도당이 포함된 YP 배지에 0.5 μg/mL aureobasidin A(Takara, Japan)와 300 μg/mL hygromycin B(Sigma-Aldrich, USA)를 첨가하였다.
본 연구에 사용한 플라스미드 목록은 Table 3에 제시하였다. 목표 DNA는 해당 primer(Table 4)와 Phusion high-fidelity DNA polymerase(New England Biolabs, USA)를 이용하여 증폭하였으며 증폭된 DNA 단편은 Gibson Assembly kit(New England Biolabs)를 사용하여 결합하였다. PSA1 및 PMI40 유전자는 각각 BamHI 및 XhoI으로 절단된 p426GPDP-CYC1T 벡터에 Gibson Assembly 방법으로 삽입하여 재조합 플라스미드를 제작하였다. 젤 정제 및 플라스미드 정제에는 Promega kit(Promega, USA)를 사용하였다.
| Plasmids | Description | References |
|---|---|---|
| p426GPDP-CYC1T | URA3, 2 μ origin, Ampr, GPD promoter, CYC1 terminator | [39] |
| p426GPDP-PSA1-CYC1T | URA3, GPDP-PSA1-CYC1T, 2 μ origin, Ampr | In this study |
| p426GPDP-PMI40-CYC1T | URA3, GPDP-PMI40-CYC1T, 2 μ origin, Ampr | In this study |
| pAUR-Cas9 | AUR1-C, CEN6, ARS4, TEF1P-Cas9-CYC1T | [40] |
| pRS42H-INT#4 | HygR, gBlock for intergenic region upstream ASF1 | [40] |
| pRS42H-INT#6 | HygR, gBlock for intergenic region upstream ATG33 | [40] |
| pRS42H-GDA1 | pRS42H, HygR, Guide RNAs targeting GDA1 | In this study |
| pRS42H-PFK26 | pRS42H, HygR, Guide RNAs targeting PFK26 | In this study |
| pRS42H-PFK27 | pRS42H, HygR, Guide RNAs targeting PFK27 | In this study |
만난 생합성 관련 유전자의 도입 및 결손에는 CRISPR-Cas9 매개 유전자 편집법을 적용하였다[24]. PSA1 및 PMI40 발현 카세트는 주형 DNA를 제작한 후, 목표 gRNA(guide RNA) 및 pAUR-Cas9을 포함하는 모균주에 LiAc(lithium acetate) 방법을 이용해 형질전환하였다[25]. 형질전환 균주는 선택배지(YPDAH, 20 g/L glucose, aureobasidin A, hygromycin B)에서 선별하였으며, 이후 항생제가 포함되지 않은 배지에서 계대배양하여 유전자 편집용 플라스미드를 제거하였다. GDA1, PFK26, PFK27 유전자 또한 Cas9 시스템을 이용하여 결손시켰다. 각 유전자의 상·하류 서열을 증폭한 뒤 중첩 PCR(overlap PCR)을 통해 주형 가닥을 제작하였고, 해당 gRNA 플라스미드와 함께 pAUR-Cas9을 보유한 효모에 도입하여 유전자 결손 균주를 구축하였다.
효모 세포벽 내 만난 함량은 산가수분해(acid hydrolysis)를 통해 생성된 만노스(mannose)를 정량하여 산출하였다. 산가수분해 조건은 효모 세포벽 다당류 분석을 위해 보고된 기존 연구를 참고하여 설정하였다[19]. 우선, 시판 효모 만난(yeast mannan, Sigma-Aldrich)을 이용해 만난–만노스 전환 표준조건을 확립하였다. 만난 용액(0–2.5 g/L)에 72%(v/v) 황산(H2SO4) 150 μL를 첨가하여 실온에서 3시간 반응시킨 뒤, 1.9 mL의 증류수를 가해 최종 황산 농도를 2 N(저농도 황산 조건)으로 조정한 후, 100℃에서 4시간 반응시켜 산가수분해를 수행하였다. 생성된 용액은 수산화바륨(Ba(OH)2) 용액으로 중화하여 침전물을 제거한 후, 상등액을 원심분리·여과하여 HPLC 분석에 사용하였다. 검출된 만노스 농도는 시판 만난 표준물질을 이용해 구축한 검량선 및 환산식을 통해 효모 세포벽 내 만난 함량으로 환산하였다.
효모 세포벽 유래 만난 정량을 위해 약 1 g의 건조세포(dry cell weight, DCW)를 회수(4,800×g, 15 min)한 후 멸균 증류수로 세척하고, 유리비드 기반 물리적 파쇄 또는 고압 균질기(high-pressure homogenization, HPH)를 이용하여 세포를 파쇄하였다. 유리비드 기반 파쇄는 0.5 mm 유리비드를 사용하여 20초 교반 후 30초 냉각 과정을 12회 반복하였다. 파쇄 후 유리비드는 TE buffer 5 mL로 5회 이상 세척하여 회수하였으며, 고압균질기(NLM 100; Ilsin Autoclave, Korea) 기반 파쇄는 120 MPa에서 5회 수행하였다. 파쇄 후 상등액을 제거한 바이오매스는 110℃ 오븐에서 하룻밤 건조하였다. 건조된 시료는 앞서 확립한 산가수분해 조건을 적용하여 만노스로 전환하였다.
생성된 만노스는 고성능액체크로마토그래피(high-performance liquid chromatography, HPLC; Shimadzu, Japan)를 이용하여 정량 분석하였다. 분석에는 굴절률 검출기(RI detector, SCL-40, Shimadzu)와 Rezex ROA-Organic Acid H⁺ 컬럼(Phenomenex, USA)을 사용하였다. 이동상으로는 5 mM 황산을 사용하였으며, 유속은 0.6 mL/min, 컬럼 온도는 60℃로 유지하였다.
효모 세포벽의 초미세 구조 관찰은 기존의 보고된 방법을 일부 수정하여 수행하였다[26]. 배양된 효모 세포를 원심분리로 회수한 후, 2.5% 글루타르알데하이드(0.1 M phosphate buffer, pH 7.2)에서 4℃, 12시간 동안 전고정하고, 1% 오스뮴 테트록사이드 용액으로 2시간 후고정하였다. 시료는 단계적으로 에탄올 탈수 후 에폭시 수지에 경화시켰다. 경화된 시료는 초박절편기(Leica EM UC7, Germany)를 이용해 약 70 nm 두께로 절단하였으며, 절편은 우라닐 아세테이트와 리드 시트레이트로 대조 염색하였다. 염색된 시료는 Bio-TEM(JEM-1400Plus, JEOL, Japan)에서 관찰하였고, 이미지는 한국기초과학 지원연구원(KBSI, 오창센터)에서 촬영하였다.
결과 및 고찰
시판 효모 만난을 0–2.5 g/L 농도로 준비한 후, 황산 조건에서 산가수분해 후 HPLC–RID로 검출된 만노스 농도를 분석하였다. 시판 효모 만난을 이용한 산가수분해–HPLC 조건의 적합성을 확인한 결과, 만난 농도 증가에 따라 생성된 만노스 농도가 선형적으로 증가하였다(R2=0.9994). 이는 본 연구에서 확립한 산가수분해 조건이 만난–만노스 전환에 대해 안정적인 반응성을 갖는다는 것을 의미한다.
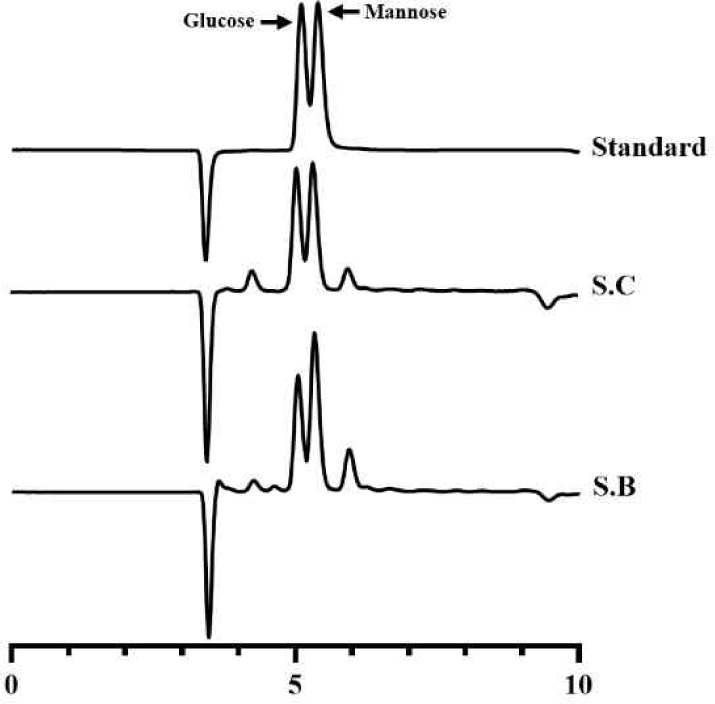
또한 본 분석법은 단당 수준에서 만노스를 정량하므로 총당법 대비 비특이적 간섭을 줄일 수 있다[6,27], 이를 검증하기 위하여 효모를 대상으로 동일한 분석을 수행하였다(Fig. 2). 그 결과, 만노스는 글루코스와 명확한 분리도를 보였으며, 효모 시료 분석 시에도 만노스 피크 영역에서 간섭 피크는 관찰되지 않았다. 실험용 효모인 S. cerevisiae D452-2를 배양 후, 건조세포중량(DCW)에 따른 만노스 함량의 변화를 정량한 결과, 세포 농도가 증가함에 따라 만노스 함량도 선형적으로 증가하였다(R2=0.9927). 이는 세포의 양에 따라 세포벽 다당류, 특히 만난의 축적이 비례적으로 증가한다는 기존 보고와 일치할 뿐만 아니라[5], 본 분석법이 실제 생물학적 시료에서도 높은 직선성과 재현성을 유지함을 보여준다. 요약하면, 시판 만난을 기반으로 구축한 표준 조건과 세포벽 유래 만난의 정량 결과가 잘 부합하였으며, 이를 통해 본 산가수분해-HPLC기반 정량법이 서로 다른 균주 및 배양 조건 간 만난 함량 비교에 신뢰성 있게 적용될 수 있음을 확인하였다.
세포벽 파쇄 방식에 따른 전처리 효율을 평가하기 위해, 유리비드 기반 파쇄와 고압균질기를 비교하였다(Fig. 3A). 산가수분해–HPLC 기반 만난 정량에서 고압균질기 처리군은 유리비드 처리군보다 높은 평균 만난 함량을 나타냈으며, 반복 측정 간 표준편차가 약 3배 이상 감소하여 분석 재현성이 크게 향상된 것으로 확인되었다.
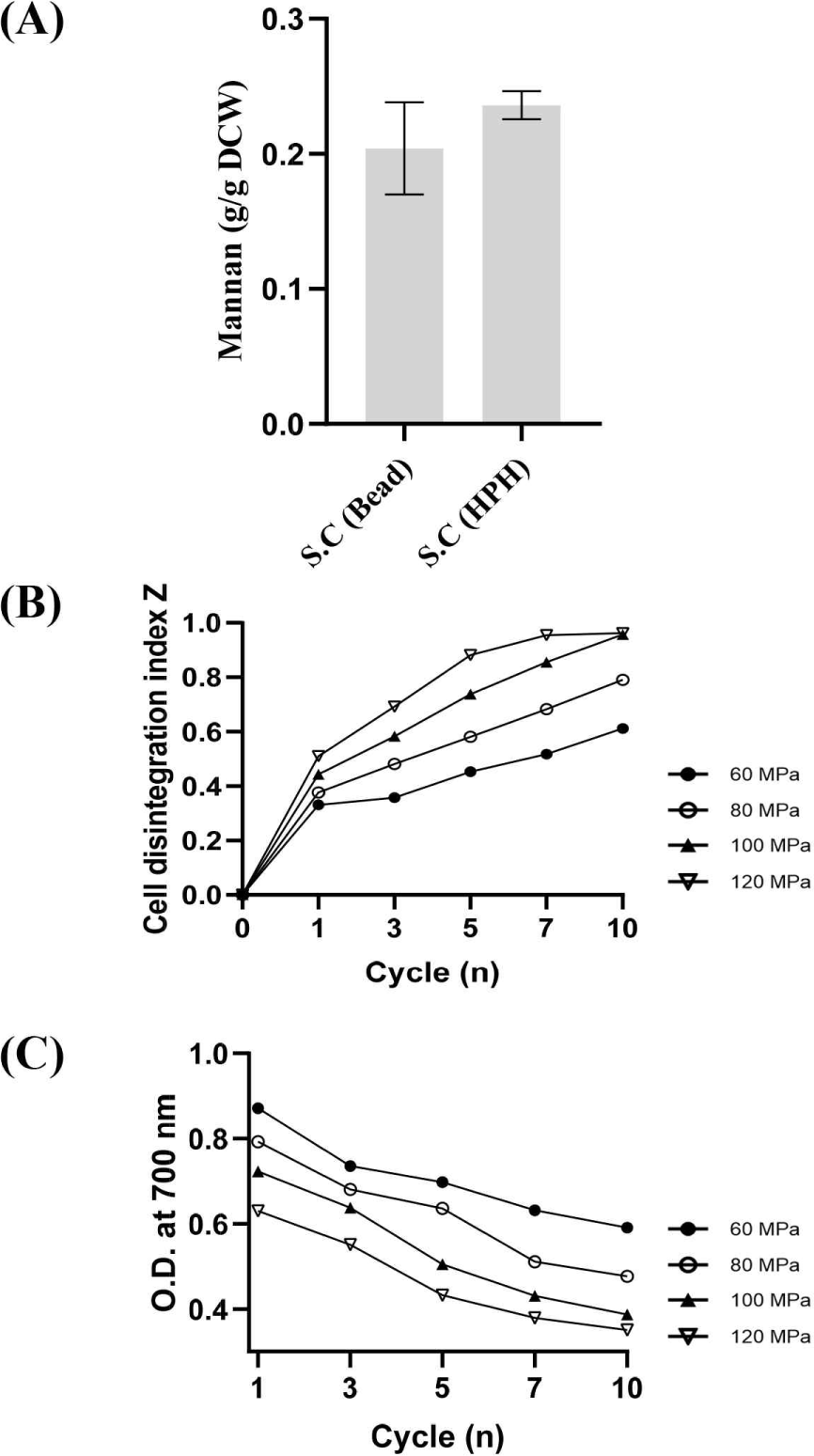
고압균질기의 파쇄 효율을 정량적으로 확인하기 위해 전기전도도 기반 세포 파괴 지수(Z-index)를 다양한 압력(60–120 MPa)과 처리 횟수(1–10 cycle) 조건에서 측정하였다(Fig. 3B). Z-index는 세포 파괴에 따라 방출되는 이온성 물질 증가를 반영하는 지표로, 0은 미파쇄 상태, 1은 완전 파쇄 상태를 의미한다[28]. 모든 압력 조건에서 초기 1–3 횟수 동안 Z-index가 급격하게 증가했으며, 이는 고압에 의해 빠른 세포벽 균열과 세포내 물질 방출이 일어났음을 시사한다. 또한 동일 횟수에서 적용 압력이 높을수록 더 높은 Z-index를 보였으며, 120 MPa에서는 10번 처리 시 Z-index가 약 0.96에 도달하여 거의 완전한 세포 파괴가 이루어졌다. 이는 고압이 강한 전단력과 압력구배를 생성하여 효율적인 세포벽 파쇄를 유도한다는 기존 보고와 일치한다[29,30].
추가적으로, 세포 파쇄 후 현탁액의 탁도를 OD700에서 측정한 결과(Fig. 3C), 압력이 증가할수록 OD700값이 감소하는 경향을 보였으며, 이는 세포벽과 세포 구성물질이 더 작은 파편으로 분해되어 빛의 산란이 감소했음을 나타낸다[30]. 이러한 현탁도 감소는 Z-index 증가 양상과 일관되며, 고압균질기가 높은 압력에서 더욱 정밀한 기계적 파쇄를 유도함을 의미한다[31].
Z-index와 OD700 두 지표는 각각 세포내 물질의 방출 정도(화학적 지표)와 세포 구조 파편화(물리적 지표)를 반영하여 세포 파괴정도를 나타내며 이를 통해 고압균질기가 유리비드 기반 파쇄에 비해 훨씬 더 강력하고 일관되게 세포벽을 파쇄함을 확인하였다. 전처리 단계의 파쇄 균일성은 산가수분해 효율과 만난 전환율, 그리고 최종 HPLC 정량 정확도를 좌우하는 핵심 요소이므로, 고압균질기의 높은 재현성은 분석 단계의 전체적인 정확도 향상에 직접적으로 기여한다. 이러한 이유로 본 연구에서는 만난 정량 플랫폼의 전처리 방식으로 고압균질기를 적용하였다.
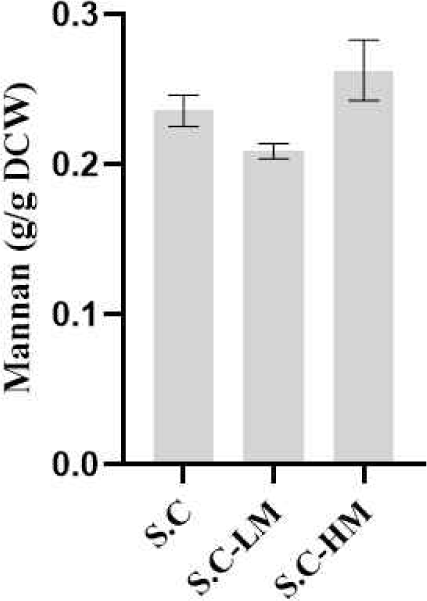
만난 함량이 상이한 균주를 확보하기 위해, 세포벽 만난 생합성에 관여하는 핵심 유전자를 표적으로 CRISPR-Cas9 시스템을 이용한 유전자 제거 및 과발현 전략을 적용하였다. 만난 저생산 균주(S.C-LM)는 GDA1 유전자를 결손하여 구축하였으며, 만난 고생산 균주(S.C-HM)은 만난 전구체인 GDP-mannose의 공급을 증가시키기 위해 PFK26과 PFK27 유전자를 파쇄하고, PMI40 및 PSA1을 과발현하도록 설계하였다.
구축한 효모의 세포벽 다당류 조성 변화를 확인하기 위해 Bio-TEM 분석을 수행하였다(Fig. 4). 야생형 S. cerevisiae는 전자밀도가 높은 내층 베타글루칸층과 이를 감싸는 외층 만난층이 명확히 구분되는 전형적인 구조를 보였다(Fig. 4A). S.C-LM 균주는 야생형 대비 외층 만난이 현저히 얇게 관찰되어, 만난 생합성 저하가 세포벽 구조 변화로 직접 반영된 것으로 확인되었다(Fig. 4B). 반면 S.C-HM 균주는 만난 층이 두껍고 균일하게 발달된 형태를 보였다(Fig. 4C).
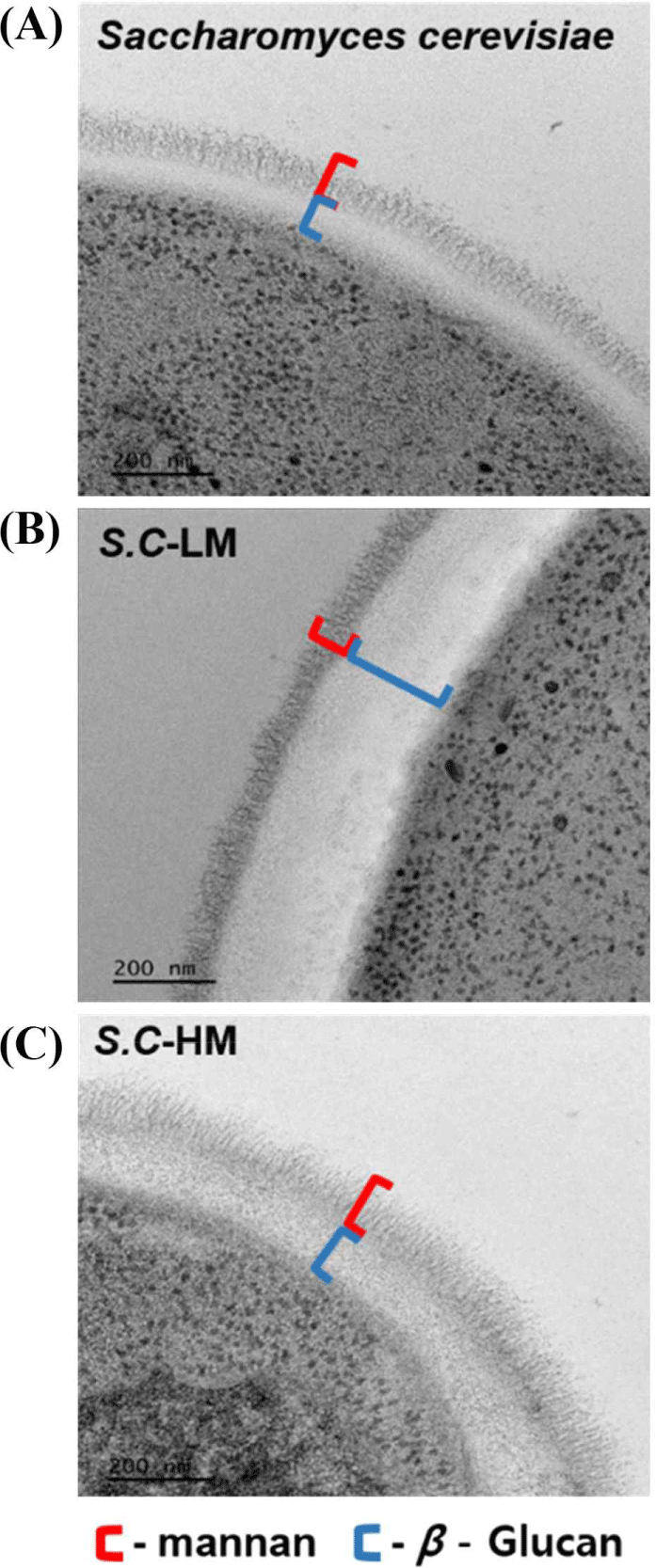
GDA1 유전자의 제거는 골지체에서의 전체적인 글리코실화 효율을 저하시켜 단백질 및 지질의 만노실화뿐 아니라 세포벽 다당류 중합을 방해하는 것으로 알려져 있다[15,16]. 이에 따라 S.C-LM 균주에서 관찰된 만난층 감소는 단순한 전구체 제한 외에 골지체 글리코실화 네트워크의 붕괴를 수반했음을 시사한다. 더불어 베타글루칸 또는 키틴 축적 증가와 같은 세포벽 보존(compensatory cell wall maintenance) 경로의 활성화 가능성도 제기되며[32,33], 이는 S.C-LM의 세포벽 미세구조 변화와도 일치한다[34,35]. S.C-HM 균주의 경우, PMI40과 PSA1 유전자의 과발현은 F6P로부터 M1P를 거쳐 GDP-mannose 로의 탄소 흐름을 증가시킨 것으로 판단된다[34]. 또한 PFK26과 PFK27 유전자의 파쇄로 인해 F6P가 F2,6bP로 전환되지 않으면서, F6P 축적이 촉진되고, 이는 결국 GDP-mannose 합성량 증가로 이어진다. 이러한 두 조절 전략의 병행은 M6P 및 M1P 축적을 통해 GDP-mannose 생합성을 극대화할 수 있으며, 해당과정 억제 시 비해당 경로로의 탄소 흐름이 증가한다는 기존 보고와도 일치한다[34]. 결과적으로, S.C-HM에서 확인된 높은 만난 축적은 현상은 단순한 유전자 과발현 효과를 넘어, 만난 생합성 경로의 병목 완화와 탄소대사 재배분이 복합적으로 작용한 결과로 해석된다.
만난 고생산 및 저생산 균주의 세포벽 만난 함량을 평가하기 위해, 본 연구에서 확립한 산가수분해–HPLC 기반 만노스 정량법을 적용하였다. 세 균주에서 동일한 건조세포중량(DCW=1 g)을 확보한 후, 황산을 첨가하여 산가수분해를 수행하였으며, 중화 후 상등액을 회수하여 HPLC-RID를 이용해 생성된 만노스 농도를 측정하였다. 측정된 만노스 값은 구축된 만노스-만난 환산식을 적용하여 최종 만난 함량을 측정하였다. 분석 결과, 저만난 균주인 S.C-LM은 야생형 효모 대비 약 0.89배 낮은 만난 함량을 나타내어 세포벽 만난이 감소했음을 확인하였다. 반면 고만난 균주 S.C-HM은 야생형 효모 대비 1.12배 높은 만난 함량을 보였으며, 이는 유전자 조작을 통한 GDP-mannose 공급 증가가 세포벽 만난 축적에 직접적으로 기여했음을 시사한다(Fig. 5).
S. boulardii는 S. cerevisiae 대비 세포벽 다당류 중 만난 비율이 높아, 장내 병원성 세균의 부착 저해 및 배출을 유도하는 대표적 프로바이오틱 효모로 알려져 있다. 본 연구에서는 산가수분해–HPLC 기반으로 구축한 만노스 정량법의 적용성을 확인하기 위해, 만난 고생산 특성을 갖는 S. boulardii를 분석 대상으로 선정하였다. 아울러 CRISPR-Cas9 시스템을 이용해 GDA1 유전자를 파쇄함으로써 만난 저생산 균주(S.B-LM)를 추가로 구축하였다.
Bio-TEM 분석에서 야생형 S. boulardii는 야생형의 S. cerevisiae(Fig. 4A)보다 더 두껍고 균일한 외층 만난층을 보여 만난 고생산 균주의 세포벽 구조를 나타냈다(Fig. 6A). 반면, GDA1 유전자가 파쇄된 S.B-LM 균주는 야생형 S. boulardii 대비 외층 만난층이 다소 얇아져 있으며(Fig. 6B), 이는 동일한 유전자 조작 전략이 두 종 모두에서 일관된 만난 감소를 유도함을 나타낸다.
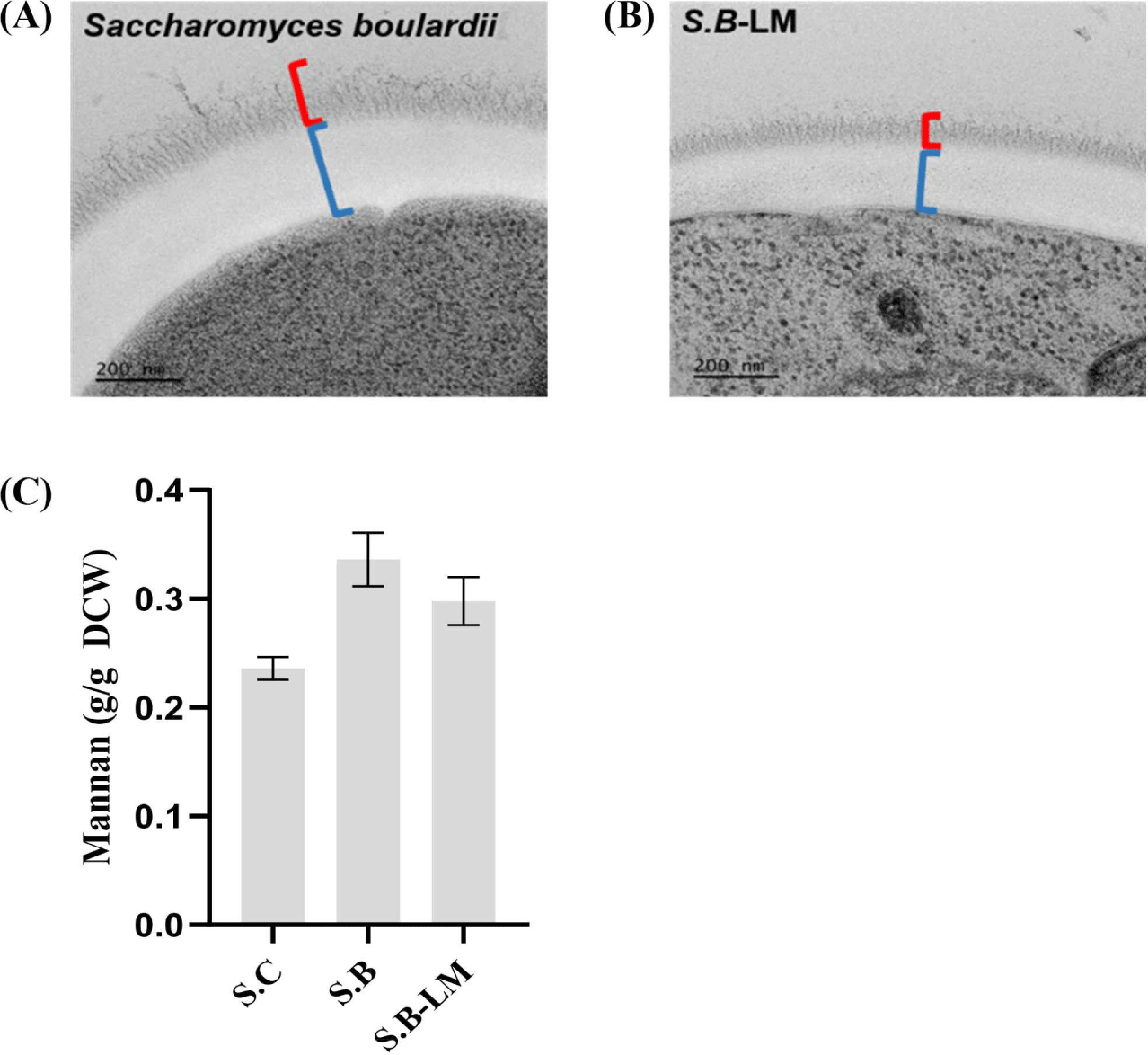
S. boulardii 야생형과 S. cerevisiae 야생형의 세포벽 만난 함량을 비교함으로써 신규 정량법의 분해능 및 종 간 적용성을 평가하였다. 정량 결과, S. boulardii 야생형에서 높은 만난 함량(0.336 g mannan/g DCW)을 나타냈으며(Fig. 6C), 이는 해당 종이 S. cerevisiae보다 세포벽 다당류 비율이 높고 고만난의 구조적 특성을 지닌다는 기존 보고와도 일치하였다[36,37]. 해당 결과는 본 정량법이 세포벽 다당류 구성이 크게 다른 종 간의 차이를 민감하게 구분할 수 있음을 보여준다.
재조합 S. boulardii 균주(S.B-LM)의 만난 함량은 야생형 S. boulardii 대비 감소하였다(Fig. 6C). 이는 S. cerevisiae의 경우와 마찬가지로 전구체 GDP-mannose 공급 제한 및 골지체 글리코실화 효율 저하로 인해 세포벽 만난 중합이 저해된 결과로 해석된다[35,36]. 이러한 정량 결과는 TEM 기반 세포벽 미세구조 변화와도 일치하였으며, S. boulardii 야생형은 두껍고 균일한 만난 층이, 저생산 균주인 S.B-LM에서는 다소 얇아진 만난층이 관찰되었다(Fig. 6B). 특히 동일한 GDA1 결손이 S. cerevisiae뿐 아니라 S. boulardii에서도 일관된 만난 감소를 유도했다는 점은 GDA1이 종 특이적 요소가 아니라 효모 전반에서 보편적으로 작용하는 핵심 조절 인자임을 시사한다[15,16].
정량 데이터와 TEM 기반 세포벽 내 만난 구성 분석이 서로 일치했다는 사실은 본 연구에서 구축한 산가수분해–HPLC 분석 플랫폼의 정확성과 생물학적 타당성을 동시에 뒷받침한다. 또한 신규 정량법이 균주 간, 종 간, 유전자 조작에 따른 세포벽 다당류 변화를 고감도로 구분한다는 점에서, 향후 기능성 효모 선발, 세포벽 다당류 대사공학, 프로바이오틱 균주 특성화 등 다양한 응용 연구에서 신뢰성 높은 만난 정량 플랫폼으로 활용될 수 있을 것으로 판단된다. 향후 세포 성장 단계별 만난 축적 양상에 대한 정량적 분석이 병행된다면, 본 분석법의 생물학적 타당성과 적용 범위가 더욱 확장될 수 있을 것으로 기대된다.
요 약
본 연구에서는 황산 기반 산가수분해와 HPLC–RID 분석을 결합한 효모 세포벽 만난 정량법을 확립하고, 대사공학적으로 구축한 다양한 효모 균주에 적용함으로써 그 활용 가능성을 평가하였다. 시판 효모 만난을 이용하여 산가수분해–HPLC 조건을 최적화한 결과, 만난 농도와 생성된 만노스 농도 사이에서 높은 상관성을 나타냈다. S. cerevisiae의 건조세포중량에 따른 만노스 농도는 선형적으로 증가하여, 세포의 양과 세포벽 만난의 선형 상관성을 보였다. 만난 저생산 균주(S.C-LM)와 만난 고생산 균주(S.C-HM)를 구축하여 산가수분해–HPLC 정량 결과, 신뢰성 있게 만난 함량을 측정할 수 있었다. 또한 해당 분석법은 프로바이오틱 효모에도 적용 가능함을 확인하였다.