




서 론
엽록체는 식물 고유의 세포소기관으로 광합성을 담당하고, 아미노산, 탄수화물, 지질, 그리고 다양한 대사산물 합성에 관여하는 등 식물 생장에 필수적인 세포소기관이다. 엽록체는 다른 세포소기관들과는 달리 이중막으로 둘러싸여 있고, 심지어 엽록체 내부에는 또 하나의 지질막으로 구성된 thylakoid가 존재한다. 이로 인해 엽록체는 3개의 지질막과 3개의 수용성 compartment로 구성되어 있는 유일한 세포소기관으로 인식된다.
엽록체는 내부공생(endosymbiosis)으로 생성된 세포소기관으로 간주되고 있고 자체의 게놈(genome)을 보유하고 있다. 하지만 엽록체를 구성하는 대부분의 단백질들은 핵(nucleus)에서 발현되고 세포질(cytosol)에 위치한 리보솜(ribosome)에서 합성된다. 단백질 합성 후에는 세포질에서 아직 잘 알려지지 않은 기작에 의해 엽록체로 이동한다(Fig. 1)[1]. 엽록체 외막 단백질은 세포질에서 외막으로 직접적으로 삽입된다[2]. 하지만 엽록체 내막, 스트로마(stroma), 그리고 thylakoid 단백질들은 엽록체 외막과 내막에 위치한 translocons at the outer and inner envelopes of chloroplasts(TOC and TIC complexes) 복합체를 통해 엽록체 내부로 수송된다[3](Fig. 1). TOC/TIC 복합체 채널을 통한 엽록체 내부로의 수송에는, 엽록체 단백질의 N-terminal 부위에 위치한 단백질 이동신호(targeting signal)인 transit peptide에 의해 이루어진다(Fig. 1)[4]. Transit peptide는 TOC 복합체의 TOC159와 TOC34 GTPase 수용체에 의해 인식된 후 TOC75 채널 단백질로 전달된다[3]. TOC75를 통과한 엽록체 단백질은 TIC 복합체의 TIC20 채널을 통해 내막을 통과한다[3]. 이 과정에서 스트로마에 존재하는 Hsp70, Hsp90, 그리고 Hsp93과 같은 ATPase 단백질의 도움으로 엽록체 내부로 단백질이 수송된다[5–7]. Transit peptide는 수송 후, 스트로마에 존재하는 단백질 분해효소인 stromal processing peptidase에 의해 절단된다[8]. Transit peptide가 제거된 mature 형태의 엽록체 단백질은 추가적인 이동신호의 존재여부 및 서열에 따라 스트로마, 내막, thylakoid 막, 또는 thylakoid lumen으로 선별(sorting) 및 이동된다. 현재까지 엽록체 외막, 스트로마, 그리고 thylakoid로 이동하는 단백질들의 이동 기작에 대해서는 상대적으로 많이 알려져 있지만, 이에 반해 엽록체 내막으로 이동하는 단백질들의 이동 및 막으로의 삽입 기작에 대해서는 연구가 미비한 측면이 있다.
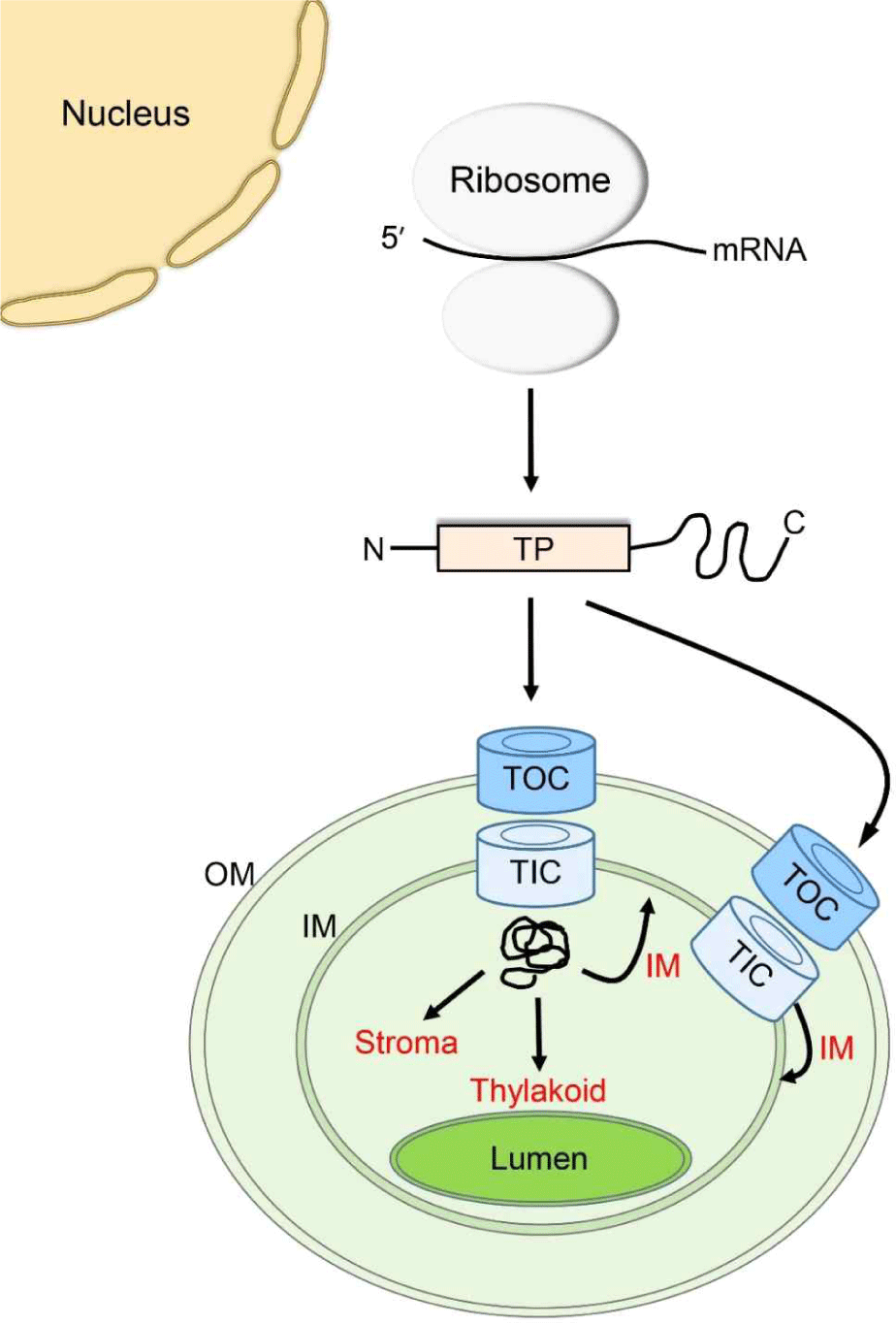
본 총설 논문에서는 상대적으로 연구가 미비한 엽록체 내막 단백질의 이동 경로(pathway)와 관련 기작에 대해 소개하고 미래의 연구 전망에 대해 고찰하고자 한다.
본 론
대부분의 엽록체 내막 단백질은 N-terminal 부위에 엽록체 이동신호인 transit peptide를 가지고 있다(Fig. 2). 다양한 transit peptide들은 일차구조에 있어서 상동성이 매우 떨어지고 심지어 consensus 서열도 존재하지 않는 특징을 가지고 있다[9]. 그럼에도 불구하고 다양한 transit peptide들은 동일한 TOC/TIC 경로를 통해 수많은 단백질들을 엽록체 내부로 수송시키는데 동일하게 사용되고 있다. 엽록체 내막 단백질들은 N-terminal transit peptide 뒤에 막 삽입에 필요한 hydrophobic transmembrane domain(TMD)을 가지고 있다. 이 TMD의 단백질 내 위치, 개수 및 삽입의 방향성(topology)은 일관되지 않고 단백질마다 고유하다(Fig. 2).
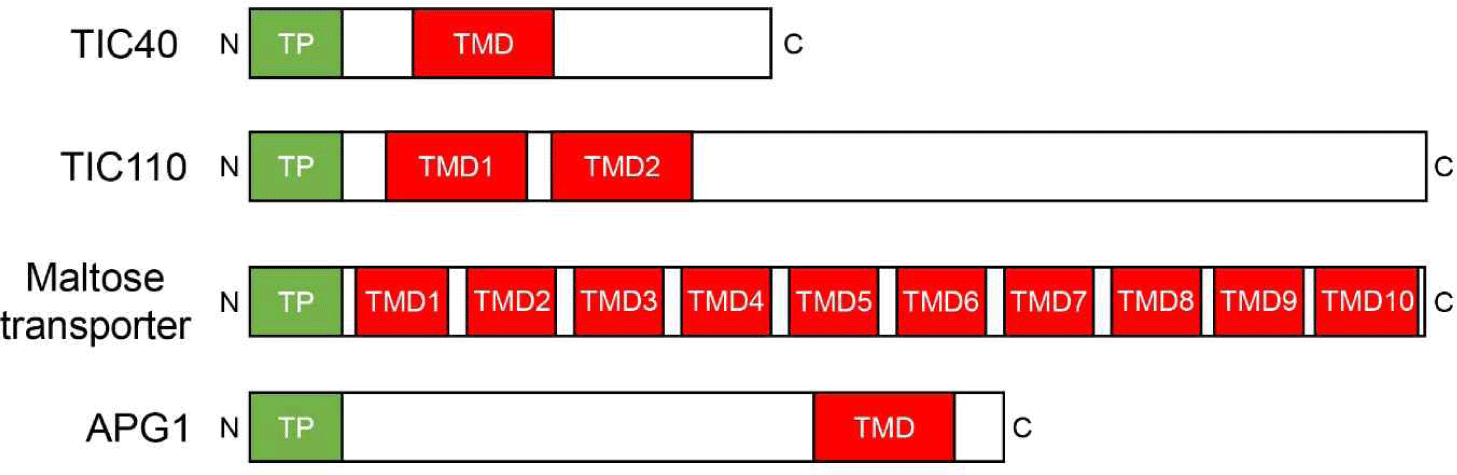
TOC/TIC 복합체를 통해 들어온 엽록체 내막 단백질은 내막에 삽입되기까지 크게 두 가지의 경로를 이용한다[1](Fig. 3). 하나는 post-import pathway이다. 예를 들어 Tic40 및 Tic110 같은 단백질은 transit peptide를 이용하여 먼저 수용성의 stroma로 수송된다. 그 후, stroma에서 엽록체 내막으로 삽입되는 경로를 이용한다[10]. 이 경로에는 내막 단백질인 Tic40이 stroma에서 엽록체 내막으로 reinsertion 과정을 통해 삽입하는데 관여한다고 보고된 바 있다[11]. 또한 엽록체 내막에 위치한 SEC2 translocase가 이 과정에 중요하게 관여한다고 알려져 있다[12,13]. 또 하나의 엽록체 내막으로의 삽입 경로는 stop-transfer pathway이다. 엽록체 내막 TIC 복합체에서 단백질 수송 채널 역할을 담당하는 단백질은 TIC20이다[14]. 그리고 stop-transfer pathway를 통해 내막으로 삽입되는 경우는, 바로 이 TIC20 채널을 통과하는 중 내막 단백질의 TMD가 내막으로 lateral하게 삽입되는 경우이다. 이 stop-transfer pathway로 삽입되는 단백질에는 APG1(albino or pale green mutant 1)가 알려져 있다. 각각의 경로를 대표하는 Tic40(post-import pathway)과 APG1(stop-transfer pathway) 서열(특히 TMD 서열)을 이용하여 다양한 돌연변이 constructs를 제작하고, in vitro import 및 분획(fractionation) 실험을 실시한 결과, Tic40과 APG1 서열의 TMD는 기본적으로 stop-transfer pathway를 이용하여 내막에 삽입되는 특성을 가진 것으로 보고되었다[15]. 그럼에도 불구하고 Tic40이 post-import pathway를 채택한 이유는 무엇일까? Tic40의 경우에는 stop-transfer pathway를 선호하는 TMD 서열과 더불어 Ser/Pro-rich 서열을 추가적으로 가지고 있는데, 바로 이 추가적인 서열이 stop-transfer 경로를 우회해서 post-import pathway를 채택하게끔 한다는 보고가 있었다[16]. 그리고 이러한 실험 결과는 post-import pathway는 stop-transfer pathway로부터 진화되었다는 점을 시사한다.
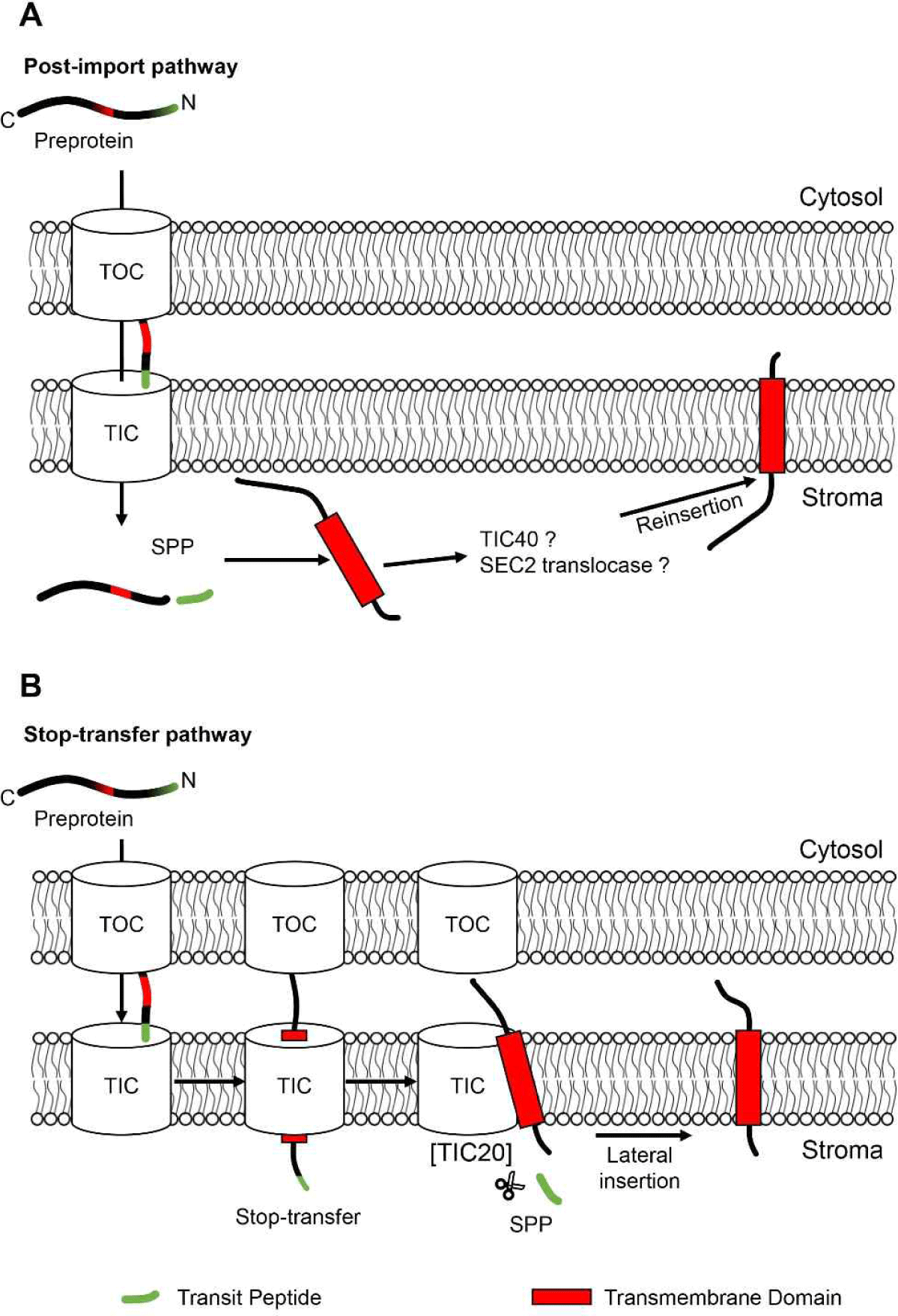
그렇다면 엽록체 내막 단백질과 thylakoid 막 단백질은 어떻게 선별되어서 각각의 막으로 삽입되는 것일까? 이 질문을 해결하기 위해서 Froehlich & Keegstra는 엽록체 내막 단백질과 thylakoid 막 단백질 서열을 이용하여 다양한 swapping 돌연변이 constructs를 제작하였다. 그 후 in vitro import 및 분획 실험을 통하여, TMD의 서열이 내막 및 thylakoid 막 단백질의 선별 및 삽입에 관여한다는 것을 밝혀냈다[17].
소수성의 TMD를 지닌 막 단백질은 세포질 같은 수용성의 환경에서는 misfolding이 일어나 단백질의 응집(aggregation) 등을 유발할 수 있고, 이는 세포에 심각한 독성을 초래할 수 있다. 엽록체 막 단백질들은 수용성의 세포질을 거쳐 엽록체로 이동해야 하는데, 이 과정에서 TMD가 세포질에서 적절히 보호되지 않으면 엽록체로의 정상적인 이동 과정에 많은 지장을 초래할 수 있다. 현재까지는 엽록체 단백질의 세포질에서의 이동 기작에 대해서 명확히 알려진 바가 없다. 추후 다양한 엽록체 단백질들과 세포질에 위치하는 chaperone 단백질 간의 상호작용, 그리고 엽록체 단백질과 세포질에서 상호작용하는 미지의 cytosolic factor들을 동정하는 과정을 통해, 막 단백질을 포함한 다양한 엽록체 단백질의 세포질에서의 이동 기작을 규명할 필요가 있다.
막 단백질의 TMD는 소수성의 지질 막에 삽입되는데 많은 경우, insertase로 기능하는 막단백질의 도움을 받아 삽입된다. 전통적으로 소포체 막으로의 단백질 삽입 기작에 대한 연구는 방대하게 이루어졌다[18]. 하지만 아직 엽록체 내막에 존재하는 insertase 단백질은 명확히 동정된 바가 없다. 앞서 언급한 SEC2 translocase의 insertase로서의 가능성을 검증하는 연구와 더불어 추가적인 insertase 내막 단백질들을 동정하는 연구가 필요할 것으로 판단된다.
마지막으로, 엽록체 내막에는 single-TMD가 아닌 다수의 TMD를 지닌 multi-pass 막 단백질들이 보고되고 있다(Fig. 2). 소포체의 경우 multi-pass 막 단백질들을 signal recognition particle 복합체가 순차적으로 TMD를 인식하여 막으로 삽입시키는 기작이 보고되었지만, 아직 다른 소기관에서는 이와 관련된 기작이 규명되지 않은 상태이다. 향후 연구를 통해, 엽록체 내막에서 다수의 TMD들을 엽록체 내막으로 삽입시키는 기작 및 TMD가 삽입되는 방향성(topology)을 결정하는 기작에 대한 연구를 수행할 필요가 있을 것으로 사료된다.
결 론
엽록체는 식물 고유의 소기관으로서 광합성을 담당하고 식물의 전주기적인 발달 및 생장에 필수 불가결한 소기관이다. 따라서 엽록체의 발달 과정은 식물의 생장 및 작물의 생산성과 깊이 연결되어 있다. 엽록체는 자체의 게놈을 가지고 있지만, 엽록체의 발달은 핵에서 발현된 수천 가지 단백질의 세포질에서의 합성, 이동, 그리고 엽록체 내부에서의 선별 기작에 의존한다. 더욱이 엽록체는 3개의 지질막 시스템을 가지고 있고 소수성의 TMD는 지질막과 상호작용하는 특징이 있는 것을 고려하면, 매우 다양한 엽록체 막 단백질들을 엽록체 내부에서 정교하게 선별하고 해당 막에 특이적으로 삽입시키는 기작은 엽록체 발달 기작을 이해하는데 매우 중요하다.
현재까지는 주로 엽록체 스트로마로 이동하는 단백질들이 TOC/TIC 복합체를 통해 수송되는 기작에 대한 연구가 이루어졌다[19]. 향후 엽록체 내막 및 thylakoid 막 단백질들의 이동 및 삽입 기작을 이해하고 구체화시키려는 다양한 시도는, 엽록체 발달 기작을 총제적으로 이해하는데 많은 도움을 줄 것으로 기대하고 궁극적으로는 기후변화의 위기 가운데 작물의 생산성을 높이는데 기여할 것으로 사료된다.