Article
신규 말벌 유인제의 등검은말벌(Vespa velutina) 포획 효능 검정
정준성1, 왕아라1, 정수연1, 김종석1, 김민지1, 황은주1, 이준길2, 이진실2, 김인선3, 김익수1,*
Efficacy Test of New Attractant for the Yellow-legged Hornet, Vespa velutina (Hymenoptera: Vespidae)
Jun Seong Jeong1, Ah Rha Wang1, Su Yeon Jeong1, Jong Seok Kim1, Min Jee Kim1, Eun Ju Hwang1, Jun Gil Lee2, Jin Sil Lee2, In Seon Kim3, Iksoo Kim1,*
Author Information & Copylight ▼
1Department of Applied Biology, Chonnam National University
2DAMOK Ecological Technology
3Department of Agricultural Chemistry, Chonnam National University
© Copyright 2018, Institute of Agricultural Science & Technology, Chonnam National University. This is an Open-Access article distributed under the terms of the
Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/4.0/) which permits
unrestricted non-commercial use, distribution, and reproduction in any
medium, provided the original work is properly cited.
Published Online: Dec 31, 2018
ABSTRACT
The yellow-legged hornet, Vespa velutina nigrithorax (Hymenoptera: Vespidae), which invaded South Korea in 2003 caused a great damage to honeybee industry. In this study, we tested the efficacy of new attractant and on-the-market-attractant in the apiaries located in Jeonnam area. As the efficacy test result, it turned out that V. velutina was the most dominant species. The newly developed attractant showed substantially high capture of V. velutina that was statistically supported (P < 0.01). On the other hand, many other insects have also been caught by the attractant, requiring further development aimed specific to V. velutina.
Keywords: Vespa velutina; Hornet; Invasive species; Attractant; Trap
서 론
국내 발생 말벌은 좀말벌(Vespa analis), 큰홑눈말벌(V. binghami), 말벌(V. crabro flavofasciata), 꼬마장수말벌(V. ducalis), 검정말벌(V. dybowskii), 장수말벌(V. mandarinia), 털보말벌(V. simillima simillima) 등 총 7종과 각 말벌(V. crabro flavofasciata)과 털보말벌(V. simillima simillima)의 아종인 등무늬말벌(V. crabro crabroniformis)과 황말벌(V. simillima xantoptera) 등 두 종의 아종이 보고되었다(Kim et al., 1994; Kim and Kim, 2011; Choi et al., 2013). 여기에 외래종인 등검은말벌(V. velutina) 을 추가하여 국내 말벌속은 총 8종, 2아종이 보고되었다(Kim et al., 2006a).
전통적으로 우리나라 양봉장에서 꿀벌을 가해하는 대표적인 말벌로는 장수말벌, 꼬마장수말벌, 좀말벌, 말벌, 털보말벌 등이 알려져 있으나(Chang et al., 1993; Jeong et al., 2007), 최근 국내 남부지방 양봉장에서 가장 우세한 말벌은 등검은말벌로 알려져 있다(Sim et al., 2014; Choi et al., 2015). 등검은말벌은 본래 아프가니스탄, 인도, 인도네시아, 중국 등 동남아시아~중국 남부 지역에 분포하였으나(van der Vecht, 1957), 국제적으로 유럽과 일본, 한국에 침입하여 확산되고 있다. 프랑스에서는 2004년 침입하여(Arca et al., 2015) 연간 80~100 km의 속도로 빠르게 확산되었으며, 현재 유럽국가에서는 스페인, 포루투갈, 이탈리아, 독일, 벨기에, 영국까지 확산되었다(López et al., 2011; Grosso-Silva and Maia, 2012; Rome et al., 2012; Federazione Apicoltori Italiani, 2013; Monceau et al., 2014; Witt, 2015; Rome et al., 2015). 일본에서는 2012년 대마도에서 처음 보고되었고(Sakai and Takahashi, 2014), 2015년 큐슈에서 발견되었다(Minoshima et al., 2015). 국내에서는 2003년 부산항을 통한 수입 목재에 의해 침입된 것으로 추측하였고, 2006년 처음으로 보고되었다(Kim et al., 2006a).
등검은말벌은 꿀벌 봉통 앞에서 제자리 비행을 하다가 봉통 밖으로 오가는 꿀벌을 공중에서 한마리씩 잡은 후 주변 나무나 건물 구조물 등에 앉아 가슴부위를 섭취한 후 나머지 부위는 버리는 행동을 보이며, 토종말벌보다 꿀벌을 사냥하는데 걸리는 시간이 짧고, 공격횟수가 많으며, 사냥 성공률이 높아 양봉 산업에 극심한 피해를 주고 있다(Jung et al., 2008; Villemant et al., 2011; Monceau et al., 2014; Choi et al., 2016). 또한 양봉장을 벗어난 민가에 대한 피해도 증가되고 있는 실정이다(YTN Science, 2016, https://www.youtube.com/watch?v=syl3WjAMQUI).
국내외적으로 등검은말벌의 방제를 위하여 유인제, 포획기, 꿀벌 개체군의 격리, 생물학적 방제(세균 및 원생생물, 곤충병원성세균, 병원성곰팡이 및 응애), 하모닉 레이더를 이용한 말벌집 탐색 등 많은 방제 방안이 보고된 바 있다(Rose et al., 1999; Kim et al., 2006b; Jung et al., 2007; Maher and Thiery, 2010; Couto et al., 2014; Milanesio et al., 2016). 무수히 날아오는 등검은말벌을 제거하기 위하여 양봉 농가에서는 배드민턴채를 이용한 포살과 트랩을 이용한 포획법에 크게 의존하고 있는 실정으로, 말벌 트랩과 유인제에 대하여 이미 다수의 연구가 진행된 바 있지만(Maher and Thiery, 2010; Couto et al., 2014; Demichelis et al., 2014), 보다 획기적인 방법이 고안되기 전까지 이들에 대한 지속적인 연구가 필요한 실정이다. 최근 프랑스에서는 등검은말벌을 효과적으로 유인할 수 있는 물질을 탐색하기 위해 벌집구성물(화분, 꿀, 꿀벌 왁스, 프로폴리스 등), 먹이원(물고기, 고기 등), 페로몬 등 자연에서 확보할 수 있는 물질과 geraniol, β-ocimene, p-xylene 등의 화학 물질의 유인 효과에 대한 Choice test 결과를 보고한 바 있다(Couto et al., 2014; Wen et al., 2017). 국내 유인제 연구의 경우, 목초, 포도주, 흑맥주 조성액 중 포도주를 사용하였을 때 등검은말벌의 유인 효과가 가장 높다고 보고된 바 있고(Sim et al., 2014), 꿀벌추출액 + 막걸리 + 당액으로 조성된 유인제에 가장 많은 개체가 유인되었다(Kang et al., 2016). 한편, 여러가지 식물 천연물(목본류, 초본류, 과일류, 해조류 등 8가지)을 이용한 등검은말벌의 유인력 비교 결과, 일정 당도 이상에서는 포획에 차이가 없는 것으로 관찰되었고, 비릿한 성분을 내는 초본류 식물에 가장 많은 등검은말벌이 포획되었다. 이에 초본류 유인제에 대한 성분분석 결과, 3-methyl-1-butanol 및 2-phenylethanol이 가장 높은 수치를 보였고, 초본류 유인제에 유일하게 acetophenone이 함유되어 있음을 보고한 바 있다(Lee, 2017).
따라서 본 연구에서는 시판 중인 ㈜다목에코텍의 말벌 유인제(이하 기존 유인제)와 이후 개선된 말벌 유인제(이하 신규 개발 유인제)에 대한 비교 실험을 통하여 신규 개발 유인제의 개선 여부를 판단하고자 하였다. 아울러 이 과정에서 축적된 양봉장 출현 말벌속(Vespa) 종의 발생양상을 정리하였다.
재료 및 방법
1. 실험장소
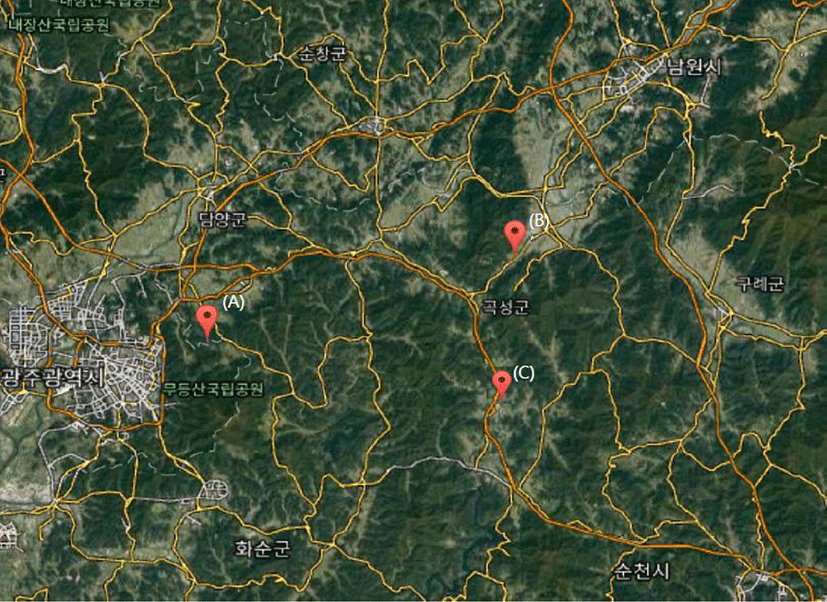
등검은말벌의 유인력 비교 실험을 위하여 2015년 말벌에 의한 피해가 컸던 담양과 곡성 각 1개소 및 2개소(담양 남면 = Damyang A, 곡성군 곡성읍 = Gokseong A, 곡성 목사동면 = Gokseong B) 등 총 3개소의 양봉장을 선정하여 ㈜다목에코텍의 포획기를 설치하였다(Fig. 1; Fig. 2). 담양 A는 남쪽으로 국립공원인 무등산과 매우 근접하고, 동쪽으로는 큰 호수가 있다. 곡성 A는 북~서쪽으로 동악산(737 m)이 뻗어 있고, 동~남쪽으로는 도로길을 따라 주로 농장이나 밭이 분포하고 있다. 양봉장 바로 앞쪽에는 도로와 체험학습장 등이 있다. 곡성 B는 주변에 과수원과 밭으로 이루어져 있고, 작은 산(220 m) 중턱에 위치하고, 북서쪽으로는 보성강이 흐르고 있다.
Fig. 1.
The Google Maps showing experimental sites (https://www.google.co.kr/maps). (A) Damyang A (Hakseon-ri 574, Nam-myeon, Damyang-gun, Jeollanam-do Province), (B) Gokseong A (Guwon-ri 63-29, Gokseong-eup, Gokseong-gun, Jeollanam-do Province), and (C) Gokseong B (Gongbuk-ri 654, Moksadong-myeon, Gokseong-gun, Jeollanam-do Province).
Download Origianl Figure
Fig. 2.
Experimental sites shown by GPS map (the upper) (NaverMaps, https://map.naver.com/)and panoramic view picture (the middle and lower). (A) Damyang A, (B) Gokseong A, and (C) Gokseong B. Red boxes in GPS Maps indicate actual experimental site.
Download Origianl Figure
2. 유인제
유인제는 기존 유인제와 신규 유인제를 사용하였다. 기존 유인제는 설탕물과 참나무 수액을 주요 성분으로 제조되었고, 신규 개발 유인제는 기존 유인제에 어성초 침지액을 추가하여 제조되었다. 유인제 실험에 사용된 트랩은 ㈜다목에코텍에서 판매중인 말벌 포획기를 사용하였다(Fig. 3).
Fig. 3.
Commercial traps used to collect Vespa including V. velutina. (A) An assembled trap, along with commodity packing box, manufactured by DAMOK Ecological Technology. (B) Entrance part, (C) Cylinder-shaped attractant container (Ø 13.5 cm), (D) Collecting bottle (56 holes, Ø 6 mm) and (E) Shield part for the prevention of captured Vespa to access to attractant solution.
Download Origianl Figure
포획기의 하단부 통에 기존 유인제와 신규 개발 유인제 각 200 mL씩을 넣고 조립 후(Fig. 3), 각 트랩간 1~5 m 정도의 거리를 두고 1~2 m 높이에 포획기를 설치하였으며, 반복구 간 거리는 20~50 m로 한 지역당 기존 유인제 및 신규 개발 유인제를 담은 트랩 총 3쌍씩을 설치하였다. 유인제 회수 시 각 유인제의 위치를 바꾸어 위치에 따른 혼란을 최소화하였다. 2016년 9월부터 11월까지 7~10일 간격으로 총 12회 조사하였다. 매 회 조사시 트랩 내 포획 개체수는 수거하고 새로운 유인제를 장착한 포획기를 설치하였다(Fig. 2).
3. 종 확인
회수한 포획기는 전남대학교 친환경농업연구소 곤충분자계통분류 및 생태학 실험실에서 포획된 개체에 대해 육안으로 분류 후 일일이 그 수를 확인하였다. 말벌속의 분류는 Kim et al. (1994)과 Jung et al. (2007) 말벌속(Vespa)의 분류키를 이용하여 분류하였다. 간략히 서술하자면, 장수말벌과 꼬마장수말벌은 다른 말벌들에 비해 크기가 크고 장수말벌은 배마디가 검은색과 주황색의 줄무늬가 반복되며, 마지막 배마디가 주황색을 나타내고, 꼬마장수말벌은 배마디에 주황색과 흑색의 줄무늬가 반복되며 5, 6, 7배마디는 흑갈색을 나타낸다. 좀말벌, 털보말벌, 말벌은 각각 배 첫번째 마디 앞가두리의 폭이 넓은 노란 띠 무늬, 물결 및 폭이 좁은 선 모양에 가깝다. 그리고 좀말벌은 등이 검고 다른 색이 없으며 털보말벌은 다른 말벌에 비해 몸 전체에 털이 많고 등면 앞, 가운데(날개 부착점) 및 뒷부분의 양 끝에 주황색 무늬가 있다. 말벌은 털보말벌에 비해 털이 적으며, 등면 앞부분과 가운데 부분(날개 부착점) 양 끝에 주황색 무늬가 있다.
등검은말벌은 전체적으로 검은색을 띠고, 다리 끝이 노랗고 배에 주황색과 검은색의 줄무늬가 있으며, 배마디 첫마디에 노란 선모양의 띠가 있고, 2, 3번째 마디에는 주황색의 넓은 띠가 있으며, 끝마디는 검은색이다. 등검은말벌 체장은 2.2~2.5 mm이며, 머리는 대부분 검은색이며, 두순, 안테나의 아랫부분, 큰 턱은 노란색을 띠고, 기부의 앞쪽만 노란색이고 나머지 머리 부분는 검은색이다(Jung et al., 2008).
말벌속의 각 종과 꿀벌(Apis mellifera)은 종 단위로 분류하였고, 말벌과(Vespidae)에서 말벌속(Veapa)과 땅벌속(Vespula)은 속 단위, 쌍살벌아과(Polistinae)는 아과 단위, 이외의 곤충은 목(Order) 단위로 분류하여 개체수를 계수하였다. 5 mm 이하의 곤충(예, 초파리류, 미소나방류 등)은 데이터에서 제외하였다.
4. 통계
신규 개발 유인제의 효능 검정을 위해 각 기존 vs. 신규 개발 유인제에 포획된 등검은말벌의 개체수를 등검은말벌의 포획 비율로 변환하여 통계처리 하였다[예, 신규 개발 유인제의 등검은말벌 포획비율 = 신규 개발 유인제의 등검은말벌 포획량 / (기존 + 신규 개발 유인제의 등검은말벌 포획량) × 100]. 포획 비율의 오차를 줄이기 위해 각 포획기에서 등검은말벌 포획량의 합이 10개체 이하일 경우와 등검은말벌 포획개체의 양 극단치는 데이터에서 제외하였다. 통계프로그램은 IBM SPSS Statistics software version 23.0 을 이용하여 t-Test를 진행하였고, 검정값을 50으로 설정하여 실시하였다.
결과 및 고찰
1. 말벌속(Vespa) 포획 양상
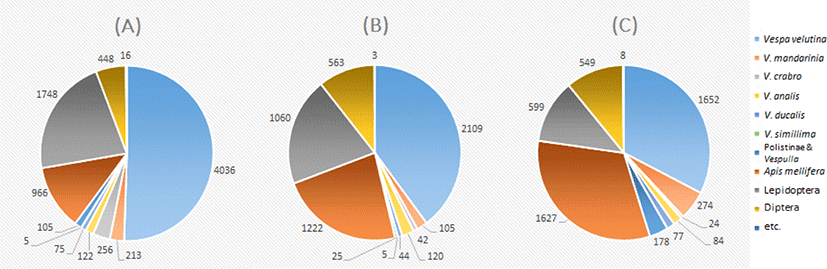
말벌속(Vespa) 포획 개체수를 비교한 결과, 세 지역 모두 등검은말벌(7,797개체, 84.3%)이 가장 많이 포획되었고, 다음으로 장수말벌(592개체, 6.4%), 좀말 벌(326개체, 3.5%), 말벌(322개체, 3.5%), 꼬마장수말벌(196개체, 2.1%), 털보말벌(10개체, 0.1%) 순이었다(Fig. 4).
Fig. 4.
Diagrams showing proportion (number) of captured insects in the three apiaries. (A) Damyang A, (B) Gokseong A, and (C) Gokseong B.
Download Origianl Figure
세 지역의 포획 개체수를 비교한 결과 말벌속내 등검은말벌의 포획비율은 담양 A, 곡성 A, 곡성 B에서 각각 85.7%, 87.0%, 78.3%로 타 말벌에 비해 가장 높게 나타났다(Fig. 4). 등검은말벌을 제외한 포획 비율을 살펴보면 담양 A는 말벌(5.4%) > 장수말벌(4.5%) > 좀말벌(2.6%) > 꼬마장수말벌(1.6%) > 털보말벌(0.1%) 의 순으로 나타났고, 곡성 A는 좀말벌(4.9%) > 장수말벌(4.3%) > 꼬마장수말벌(1.8%) > 말벌(1.6%) > 털보말벌(0.2%)의 순으로, 곡성 B는 장수말벌(13.0%) > 좀말벌(4.0%) > 꼬마장수말벌(3.6%) > 말벌(1.1%) 순으로 나타났으며, 곡성 B에서는 털보말벌이 포획되지 않았다. 이상의 결과를 살펴보면, 등검은말벌의 우점도가 가장 높았으며, 등검은말벌을 제외한 말벌들은 그 비율이 매우 낮음을 확인하였고, 지역간 발생하는 말벌 종과 빈도의 차이가 있음을 확인하였다. Sim et al. (2014)의 2014년 4월부터 7월까지 곡성에서 포획된 말벌속 종들의 포획 비율과 비교해 보았을 때(등검은말벌 11.4%, 장수말벌 31.4%, 꼬마장수말벌 11.9%, 털보말벌 24.9%, 검정말벌 1.6%, 좀말벌 2.2%, 말벌 16.8%), 현재 곡성의 등검은말벌 포획 비율이 압도적으로 높아진 것을 확인하였다. 그리고 등검은말벌을 제외한 나머지 말벌속의 개체 비율을 비교한 결과, 털보말벌, 검정말벌, 말벌의 비율이 감소하였고, 장수말벌 비율은 곡성 A에서는 감소, 곡성 B에서는 상승하였다. 꼬마장수말벌은 소폭 상승하였으며, 좀말벌의 비율은 크게 증가하였다(Table 1).
Table 1.
Trapping ratio of Vespa species in Gokseong regions excluding V. velutina (%)
|
|
V. mandarinia
|
V. ducalis
|
V. simillima
|
V. dybowskii
|
V. analis
|
V. crabro
|
| Gokseong by Sim et al. (2014) |
35.4 |
13.4 |
28.1 |
1.8 |
2.5 |
19.0 |
| Gokseong A (This study) |
33.2 |
13.9 |
1.6 |
0.0 |
38.0 |
13.3 |
| Gokseong B (This study) |
59.7 |
16.8 |
0.0 |
0.0 |
18.3 |
5.2 |
Download Excel Table
2. 시기별 말벌속 포획양상
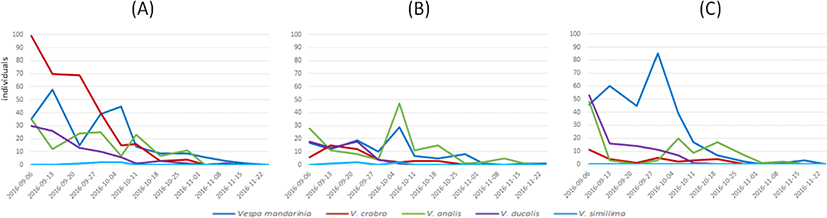
시기별 말벌속 종의 포획 양상을 살펴 본 결과, 장수말벌은 9월 초 포획되는 개체수가 감소하다가 10월 초 다소 증가했으며, 중순 이후 감소하는 추세를 나타냈다. 꼬마장수말벌의 경우, 9월 초에 가장 높은 개체수를 나타났으며, 이후 감소하는 추세를 나타냈다. 말벌의 경우, 9월 초 가장 많이 포획되었고, 좀말벌의 경우 10월 초에 가장 많이 포획되었다. 털보말벌의 경우, 포획된 개체가 적지만 9월 말~10월 초에 가장 많이 포획되었다(Fig. 5). 등검은말벌은 세 지역 모두 9월 초부터 포획 량이 증가하였고, 10월 초에 가장 많은 수가 포획되었고, 이후 개체수가 급감하는 것을 확인하였다(Fig. 6).
Fig. 5.
Trapping patterns of five Vespa species during surveyed period (excluding V. velutina) in three places of apiaries. (A) Damyang A, (B) Gokseong A, and (C) Gokseong B.
Download Origianl Figure
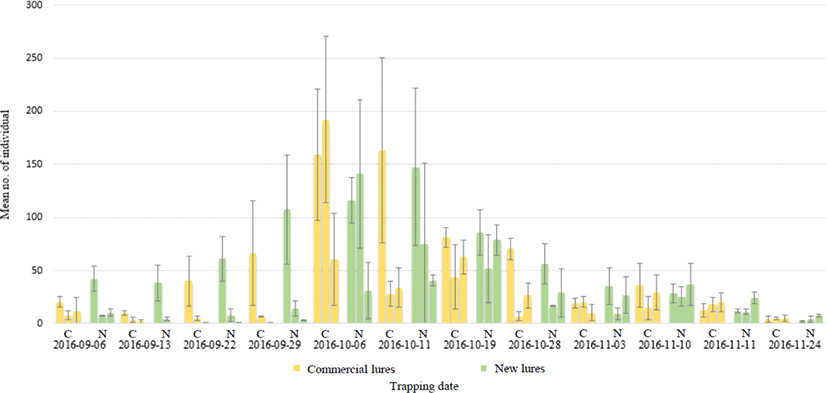
Fig. 6.
Trapping pattern for Vespa velutina between commercial and new attractants. C (Orange bar) and N (Green bar) mean each commercial and new attractant. Error bar indicate standard deviation.First, second, third bar indicate mean number of V. velutina captured each at Damyang A, Gokseong A, and Gokseong B.
Download Origianl Figure
3. 신규 개발 유인제 포획력 검정
두 유인제에 의한 전체 포획 개체를 살펴보면 신규 개발 유인제에 더 많은 등검은말벌 [기존 = 3,786 (40.1%), 신규 = 4,011 (53.9%)]이 포획되었다. 반면, 장수말벌 [기존 = 315(3.3%), 신규 = 277 (3.1%)], 말벌 [기존 = 179 (1.9%), 신규 = 143 (1.6%)], 좀말벌 [기존 = 194 (2.1%), 신규 = 132 (1.5%)], 꼬마장수말벌 [기존 = 124 (1.3%), 신규 = 72 (0.8%)], 털보말벌 [기존 = 6 (0.1%), 신규 = 4 (0.0%)], 파리목 [기존 = 903 (9.6%), 신규 = 657 (7.4%)], 나비목 [기존 = 2,042 (21.6%), 신규 = 1,365 (15.3%)]은 신규 개발 유인제보다 기존 유인제에 더 많이 포획되었다. 꿀벌 또한 등검은말벌처럼 신규 개발 유인제에 더 많이 포획되었다[기존 = 1,676 (17.8%), 신규 = 2,139 (24.0%)] (Table 2).
Table 2.
Comparison of the numbers (ratio) of Vespa species trapped in commercial attractant (C) and new attractant (N)
|
|
|
Damyang A |
Gokseong A |
Gokseong B |
Tatal |
|
V. velutina
|
C |
1955 (47.3) |
1044 (37.4) |
787 (31.3) |
3786 (40.1) |
| N |
2081 (53.9) |
1065 (42.5) |
865 (33.8) |
4011 (44.9) |
|
V. mandarinia
|
C |
106 (2.6) |
56 (2.0) |
153 (6.1) |
315 (3.3) |
| N |
107 (2.8) |
49 (2.0) |
121 (4.7) |
277 (3.1) |
|
V. crabro
|
C |
143 (3.5) |
22 (0.8) |
14 (0.6) |
179 (1.9) |
| N |
113 (2.9) |
20 (0.8) |
10 (0.4) |
143 (1.6) |
|
V. analis
|
C |
76 (1.8) |
66 (2.4) |
52 (2.1) |
194 (2.1) |
| N |
46 (1.2) |
54 (2.2) |
32 (1.2) |
132 (1.5) |
|
V. ducalis
|
C |
45 (1.1) |
27 (1.0) |
52 (2.1) |
124 (1.3) |
| N |
30 (0.8) |
17 (0.7) |
25 (1.0) |
72 (0.8) |
|
V. simillima
|
C |
3 (0.1) |
3 (0.1) |
0 (0) |
6 (0.1) |
| N |
2 (0.1) |
2 (0.1) |
0 (0) |
4 (0) |
| Vespula & Polistinae |
C |
53 (1.3) |
10 (0.4) |
128 (5.1) |
191 (2) |
| N |
52 (1.3) |
15 (0.6) |
50 (2.0) |
117 (1.3) |
|
Apis mellifera
|
C |
435 (10.5) |
540 (19.3) |
701 (27.9) |
1676 (17.8) |
| N |
531 (13.8) |
682 (27.2) |
926 (36.2) |
2139 (24) |
| Lepidoptera |
C |
1013 (24.5) |
702 (25.1) |
327 (13) |
2042 (21.6) |
| N |
735 (19.0) |
358 (14.3) |
272 (10.6) |
1365 (15.3) |
| Diptera |
C |
290 (7.0) |
319 (11.4) |
294 (11.7) |
903 (9.6) |
| N |
158 (4.1) |
244 (9.7) |
255 (10.0) |
657 (7.4) |
| Others1 |
C |
11 (0.3) |
3 (0.1) |
3 (0.1) |
17 (0.2) |
| N |
5 (0.1) |
0 (0) |
5 (0.2) |
10 (0.1) |
Download Excel Table
기존 유인제 대비 신규 개발 유인제 포획량에 대한 통계적 분석 결과, 담양 A는 56.1% (t = 2.195, P < 0.05), 곡성 A는 54.9% (t = 1.791, P < 0.05)로 신규 개발 유인제에 통계적으로 유의하게 더 많은 등검은말벌이 포획되었고, 곡성 B는 54.6% (t = 1.436, P = 0.08)로 통계적으로 유의하지는 않지만 유의확률 0.05와 근접한 수치를 나타내며, 신규 유인제에 더 많은 등검은말벌이 포획되었다, 지역에 관계없이 위의 세 지역에 등검은말벌 포획 비율을 비교해 보았을 때 신규 유인제가 55.22% (P < 0.005)로 통계적으로 유의하게 신규 유인제가 더 많은 포획량을 보였다(data not shown). 이로 미루어 신규 개발 유인제가 기존 유인제보다 등검은말벌 특이적으로 개선되었음을 알 수 있었다(Fig. 6; Table 2). 그러나 꿀벌 역시 더 많은 개체가 신규 개발 유인제에 포획되었으므로 이에 대한 개선이 필요한 실정이다. Demichelis et al. (2014)에 따르면, 맥주, 민트, 당 성분이 들어있는 유인제[suger (4%), honey (4%)] 중 당 성분이 들어있는 유인제를 사용하였을 때와 포획기 뚜껑의 색상을 노란 뚜껑의 트랩으로 사용하였을 때 꿀벌이 가장 많이 포획되었다고 보고되었다. 또한 Monceau et al. (2013)에 따르면 먹이 유인제를 사용하였을 때 말벌 이외의 다른 곤충이 포획되는 심각한 부작용을 일으킬 수 있다고 보고되었다. 따라서 국내외적으로 보다 말벌 특이적인 유인제 개발이 필요한 실정이다.
4. 등검은말벌 수벌 포획 양상
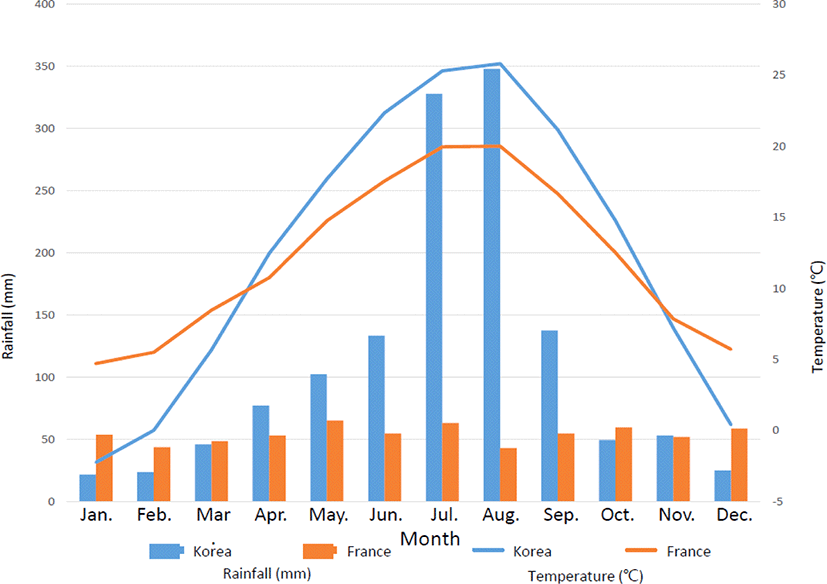
등검은말벌 수벌은 9~10월 초까지 낮은 개체수를 유지하다가 10월 중순쯤부터 개체수가 증가하기 시작하였고, 11월 초에 가장 많은 개체수를 기록하였다가 이후 감소하고 일벌(여왕벌) 대비 포획 비율 역시 11월 초가 가장 높았다(Table 3). 프랑스의 연구결과, 등검은말벌은 가을철에 생식, 분산에 집중하는 시기이고, 9월 중순에서 11월 말까지 수벌 생산이 3배 더 증가한다고 보고된 바 있고(Monceau et al., 2014), 국내의 경우 11월에 생식 활동이 가능한 수벌과 여왕벌을 가진 성숙된 colony를 갖는다고 보고되었다(Choi et al., 2012). 이를 바탕으로 볼 때 11월에 수벌이 가장 많이 잡힌 이유는 9월부터 수벌의 생산이 증가되고 생식을 위해 수벌이 봉군를 떠나 분산되면서 포획량이 증가된 것으로 추정된다. 프랑스는 지중해성 기후인 반면, 한국은 계절풍 기후로 프랑스가 한국에 비해 연교차가 작지만 두 국가 모두 7월과 8월이 최난월(최난월 기온 평균 : 한국은 25°C, 프랑스는 18°C)을 가지고 이후 온도가 떨어지는 것으로 알려져 있다. 그러므로 등검은말벌 수벌의 11월 개체수 증가는 기온과 밀접한 관련이 있는 것으로 추정된다(Fig. 7; Table 3).
Table 3.
Trapping pattern of Vespa velutina workers, queens and males (%)
| Sex |
September |
October |
November |
| 6 |
13 |
22 |
29 |
6 |
11 |
19 |
28 |
3 |
10 |
17 |
24 |
| Workers and queens |
296 (98.3) |
171 (99.4) |
336 (98.2) |
589 (99.0) |
2099 (99.9) |
1441 (98.5) |
1162 (95.9) |
580 (94.0) |
297 (81.6) |
447 (87.0) |
271 (91.9) |
72 (91.7) |
| Male |
5 (1.7) |
1 (0.6) |
6 (1.8) |
6 (1.0) |
2 (0.1) |
22 (1.5) |
50 (4.1) |
37 (6.0) |
67 (18.4) |
67 (13.0) |
24 (8.1) |
6 (8.3) |
Download Excel Table
요약하자면 외래종인 등검은말벌은 최근 전남지방에서 빠르게 적응, 확산하였다. 현재 양봉장에서 출현하는 말벌속의 발생 비율이 크게 바뀌어 등검은말벌이 가장 높은 우점도를 나타냈고, 토종말벌들의 비율은 큰 폭으로 감소한 것으로 나타났다. 이러한 등검 은말벌의 방제를 위해 기존 유인제를 개선한 신규 개발 유인제를 전남 곡성 1개소, 담양 2개소 양봉장에서 유인 효능에 대한 검정 결과, 신규 개발 유인제가 등검은말벌 특이적으로 효능이 다소 향상되었고, 이는 통계적으로 유의하였다.
요 약
본 실험에서는 등검은말벌(V. velutina) 유인제의 포획 효능을 알아보고, 등검은말벌의 발생양상을 조사하기 위해 실시하였다. 국내 발생 10종의 Vespa 속 말벌 중 등검은말벌(7,797개체)이 가장 많이 포획되었고, 다음으로 장수말벌(592개체), 좀말벌(326개체), 말벌(322개체), 꼬마장수말벌(196개체), 털보말벌(10개체) 순으로 포획되어 등검은말벌이 가장 높은 빈도로 발생함을 확인하였다. 두 유인제의 포획력 검정 결과 신규 개발 유인제가 통계적으로 유의하게(P < 0.01) 등검은말벌의 포획력이 높음을 보였고, 반면 장수말벌, 말벌, 좀말벌, 꼬마장수말벌, 털보말벌, 파리목, 나비목은 신규 개발 유인제보다 기존 유인제에서 더 많이 포획되는 점으로 미루어 신규개발 유인제가 다소 등검은말벌 특이적임을 확인하였다. 한편, 등검은말벌 수벌은 9~10월 초까지 낮은 개체수를 유지하다 10월 중순쯤부터 개체수가 증가하기 시작하였고, 11월 초에 가장 많은 개체수를 기록하다 이후 감소하였다. 등검은말벌 일벌은 9월부터 10월 초까지 개체수가 증가한 가운데 10월 초부터 중순까지 가장 많은 개체수를 기록하였고, 이후 감소하였다.
감사의 글
본 결과물은 농림축산식품부의 재원으로 농림식품기술기획평가원의 농생명산업기술개발사업의 지원을 받아 연구되었습니다 [과제번호:316038-3].
참고문헌
Arca, M., Mougel, F., Guillemaud, T., Dupas, S., Rome, Q., Perrard, A., Muller, F., Fossoud, A., Capdevielle-Dulac, C., Torres-Leguizamon, M., Chen, X. X., Tan, J. L., Jung, C., Villemant, C., Arnold, G. and Silvain, J.-F. 2015. Reconstructing the invasion and the demographic history of the yellow-legged hornet,
Vespa velutina, in Europe. Biological Invasions 17:2357-2371. .


Chang, Y. D., Lee, M. Y., Yim, Y. H. and Youn, Y. N. 1993. Species and visiting patterns of wasps (Hymenoptera: Vespoidea) in apiary. Korean Journal of Apiculture 8:22-28..
Choi, M. B., Martin, S. J. and Lee, J. W. 2012. Distribution, spread, and impact of the invasive hornet
Vespa velutina in South Korea. Journal of Asia-Pacific Entomology 15:473-477. .
Choi, M. B., Kim, J. K. and Lee, J. W. 2013. Checklist and distribution of Korean Vespidae revisited. Korea Journal of Applied Entomology 52: 85-91. .
Choi, M. B. and Kwon, O. 2016. 양봉장에서 등검은말벌의 사냥 시간, 공격 횟수 및 성공율 분석. Korean Society of Applied Entomology 4:48-48..
Choi, Y. S., Lee, M. L., Lee, M. Y., Kim, H. K., Yoon, M. Y. and Kang, A. R. 2015. Trapping of
Vespa velutina nigrithorax Buysson (Hymenoptera: Vespidae) queen near apiaries honeybee comb extract in south region of Korea. Journal of Apiculture 30:281-285. .
Demichelis, S., Manino, A., Minuto, H., Mariotti, M. and Porporato, M. 2014. Social wasp trapping in north west Italy: Comparison of different bait-traps and first detection of
Vespa velutina. Bulletin of Insectology 67:307-317..
Grosso-Silva, J. M. and Maia, M. 2012.
Vespa velutina Lepeletier, 1836 (Hymenoptera, Vespidae), new species for Portugal. Arquivos Entomoloxicos 6:53-54..
Jung, C., Kang, M. S., Kim, D. W. and Lee, H. S. 2007. Vespid wasps (Hymenoptera) occurring around apiaries in Andong, Korea. Korea Journal of Apiculture 22:53-62..
Jung, C., Kim, D. W., Lee, H. S. and Back, H. 2008. Some biological characteristics of a new honeybee pest,
Vespa velutina nigrithorax Buyssan 1905 (Hymenoptera: Vespidae). Korean Journal of Apiculture 24:61-65..
Kang, E. J., Lee, M. L., Lee, M. Y., Kim, H. K. and Choi, Y. S. 2016. Attractive effect using honeybee extraction against
Vespa velutina nigrithorax Buysson (Hymenoptera: Vespidae) queen. Journal of Apiculture 31:195-199. .
Kim, J. K., Moon, T. Y. and Yoon, I. B. 1994. Systematics of Vespine Wasps from Korea, I. Genus
Vespa Linnaeus (Vespidae, Hymenoptera). Korean Journal of Entomology 24:107-115..
Kim, J. K., Choi, M. B. and Moon, T. Y. 2006a. Occurrence of
Vespa velutina Lepeletier from Korea, and a revised key for Korean
Vespa species (Hymenoptera: Vespidae). Entomological Research 36:112-115. .
Kim, Y. S., Lee, M. Y., Lee, M. L. and Nam, S. H. 2006b. Development of natural luring liquid against the wasps inflicting honey bees. Korean Journal of Apiculture 21:37-42..
Kim, J. K. and Kim, I. K. 2011. Discovery of
Vespa binghami (Vespidae: Hymenoptera). Korea Journal of Entomology 24:107-115. .
Lee, J. S. 2017. Screening of attractant and developing of trap for the yellow-legged, hornet
Vespa velutina. Master’s degree thesis Chonnam National University..
López, S., González, M. and Goldarazena, A. 2011.
Vespa velutina Lepeletier, 1836 (Hymenoptera, Vespidae): First records in Iberian Peninsula. EPPO Bulletin / Bulletin OEPP 41:439-441. .
Maher, N. and Thiery, D. 2010. Comparison of trap design against the yellow-legged hornet
Vespa velutina. Apimondia (Presentation material)..
Monceau, K., Maher, N., Bonnard, O. and Thiery, D. 2013. Predation pressure dynamics study of the recently introduced honeybee killer
Vespa velutina: learning from the enemy. Apidologie 44:209-221. .
Monceau, K. and Bonnard, O. 2014.
Vespa velutina: a new invasive predator of honeybees in Europe. Journal of Pest Science 87:1-16. .
Minoshima, Y. N., Yamane, S. and Ueno, T. 2015. An invasive alien hornet,
Vespa velutina nigrithorax du Buysson (Hymenoptera, Vespidae), found in Kitakyushu, Kyushu Island: A first record of the species from mainland Japan. Japanese Journal of Systematic Entomology 21:259-261..
Rome, Q., Muller, F. and Villemant, C. 2012. Expansion en 2011 de
Vespa velutina Lepeletier en Europe (Hym., Vespidae). Bulletin de la Société entomologique de France 117:114..
Rome, Q., Muller, F. J., Touret-Alby, A., Darrouzet, E., Perrard, A. and Villemant, C. 2015. Caste differentiation and seasonal changes in
Vespa velutina (Hym.: Vespidae) colonies in its introduced range. Journal of Applied Entomology 139:771-782. .
Rose, E. A. F., Harris, R. J. and Glare, T. R. 1999. Possible pathogens of social wasps (Hymenoptera: Vespidae) and their potential as biological control agents. New Zealand Journal of Zoology 26:179-190. .
Sakai, Y. and Takahashi, J. 2014. Discovery of a worker of
Vespa velutina (Hymenoptera: Vespidae) from Tsuhima Island, Japan (Japanese with English summary). Japanese Journal of Applied Entomology and Zoology 17:32-36..
Sim, H. S., Lee, M. L., Choi, Y. S., Kim, H. G., Hong, I. P., Woo, S. O., Byeon, K. H. and Lee, M. Y. 2014. Pattern of emergence of
Vespa velutina nigrithorax Buysson (Hymenoptera: Vespidae) on spring in south part of Korea. Journal of Apiculture 29:353-358. .
Van Der Vecht, J. 1957. The Vespinae of the Indo-Malayan and Papuan areas (Hymenoptera: Vespidae). Zoologische Verhandelingen 34:1-82..
Villemant, C., Muller, F., Haoubois, S., Perrard, A., Darrouzet, E. and Rome, Q. 2011. Predicting the invasion risk by the alien bee-hawking yellow-legged hornet
Vespa velutina nigrithorax across Europe and other continents with Niche models. Biological Conservation 144:2142-2150. .
Wen, P., Cheng, Y. N., Dong, S. H., Wang, Z. W., Tan, K. and Nieh, J. C. 2017. The sex pheromone of a globally invasive honey bee predator, the Asian eusocial hornet,
Vespa velutina. Scientific Reports 7:12956. .
Witt, R. 2015 Erstfund eines Nestes der Asiatischen Hornisse
Vespa velutina Lepeletier, 1838 in Deutschland und Details zum Nestbau (Hymenoptera, Vespinae). Ampulex 7:42-53..